Материал: Жукова Л.А. (отв. ред.) Онтогенетический атлас лекарственных растений. Том VII

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Таблица 4 |
|
|
|
|
Биометрические показатели особей А. zingeri разных уровней жизненности |
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Морфологи- |
|
v |
|
|
g1 |
|
|
g2 |
|
|
g3 |
|
|
Ss |
|
|
ческие |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
II |
III |
I |
II |
III |
I |
II |
III |
I |
II |
III |
I |
II |
III |
|
|
признаки |
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Высота |
23,5 |
15,0 |
10,1 |
32,3 |
22,5 |
14,4 |
37,8 |
25,2 |
17,3 |
35,7 |
27,4 |
18,1 |
27,0 |
17,6 |
10,3 |
|
годичного |
|||||||||||||||
|
±0,7 |
±0,5 |
±0,4 |
±0,7 |
±0,6 |
±0,8 |
±0,5 |
±0,3 |
±0,4 |
±0,9 |
±0,6 |
±0,4 |
±0,7 |
±0,3 |
±0,4 |
|
|
побега, см |
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Длина листа, |
5,3 |
3,8 |
2,9 |
5,5 |
4,1 |
3,1 |
6,5 |
4,5 |
2,9 |
5,5 |
5,0 |
2,7 |
5,3 |
4,0 |
3,0 |
|
см |
±0,3 |
±0,5 |
±0,2 |
±0,5 |
±0,3 |
±0,4 |
±0,5 |
±0,3 |
±0,3 |
±0,4 |
±0,3 |
±0,8 |
±0,7 |
±0,3 |
±0,5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Ширина |
1,0 |
0,5 |
0,3 |
1,3 |
1,0 |
0,4 |
1,6 |
1,3 |
0,7 |
1,4 |
0,9 |
0,4 |
1,2 |
0,8 |
0,6 |
|
листа, см |
±0,3 |
±0,2 |
±0,1 |
±0,2 |
±0,1 |
±0,3 |
±0,3 |
±0,3 |
±0,2 |
±0,1 |
±0,1 |
±0,2 |
±0,2 |
±0,1 |
±0,2 |
86 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Среднее коли- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
чество генера- |
– |
– |
– |
3,4 |
2,9 |
1,5 |
27,8 |
15,3 |
10,7 |
8,9 |
5,7 |
3,6 |
– |
– |
– |
|
тивных побе- |
±0,6 |
±0,3 |
±0,7 |
±0,4 |
±0,5 |
±0,6 |
±0,3 |
±0,5 |
±0,2 |
||||||
|
|
|
|
|
|
|
||||||||||
|
гов, шт. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Среднее число |
|
|
|
13,9 |
10,8 |
7,2 |
23,6 |
14,2 |
10,9 |
18,2 |
12,4 |
10,1 |
|
|
|
|
цветков на |
– |
– |
– |
– |
– |
– |
|||||||||
|
±0,3 |
±0,5 |
±0,6 |
±0,8 |
±0,4 |
±0,5 |
±0,3 |
±0,6 |
±0,5 |
|||||||
|
побеге, шт. |
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Среднее число |
|
|
|
8,2 |
5,3 |
1,5 |
19,1 |
11,4 |
5,9 |
15,3 |
7,9 |
7,3 |
|
|
|
|
плодов на по- |
– |
– |
– |
– |
– |
– |
|||||||||
|
±0,5 |
±0,3 |
±0,6 |
±0,2 |
±0,3 |
±0,5 |
±0,4 |
±0,5 |
±0,2 |
|||||||
|
беге, шт. |
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ИММАТУРНЫЕ растения характеризуются появлением, наряду с тройчатыми, непарноперистосложных листьев в количестве 4–5 штук, шириной 10–15 мм, длиной 30–40 мм, состоящих из 2–3 пар листочков. В пазухах листьев закладываются почки. Стебель высотой 9–13 см, не одревесневший, в диаметре достигает 1–1,5 мм. Главный корень длиной 12–14 см. Продолжительность состояния 1–2 года.
ВИРГИНИЛЬНЫЕ особи характеризуются переходом от моноподиального роста к симподиальному, что наблюдается на втором (третьем) году жизни. Происходит нарастание вегетативной массы растения. Растения достигают высоты 10–20 см. Количество побегов увеличивается до 5–8. Количество листьев до 20 штук, длиной до 5 см, шириной до 1 см. В природе продолжительность состояния до 5 лет, при интродукции до года.
Далее растение вступает в генеративный период своего развития. Впервые А. zingeri зацветает в условиях интродукции на 2–3 год, в естественных условиях на 3–5 (7) год.
МОЛОДЫЕ ГЕНЕРАТИВНЫЕ особи имеют 1 (3) генеративный неспециализированный побег. Он имеет как бы две сферы – вегетативную (нижняя половина побега с почками возобновления) и генеративную (занимает верхнюю половину побега). Число цветков в соцветии до 12. Вегетативная часть представлена 4–6 побегами, одревесневшими на 1–2 см. Листьев от 3 до 30 штук, длиной до 5 см, шир иной до 1,5 см. Состарившись, после плодоношения генеративный побег отмирает, оставляя базальную часть в прирост куста. Формируется каудекс диаметром 2–3 мм. Продолжительность состояния 3(5)–5(7) лет.
СРЕДНЕВОЗРАСТНЫЕ ГЕНЕРАТИВНЫЕ растения достигают в высоту
20–30 (40) см. Наблюдается переход большинства побегов из вегетативных в генеративные. Характерно пышное развитие генеративных побегов в количестве 10 (25) штук. Цветков в головчатой кисти до 20 штук. В наиболее сухие годы преобладают вегетативные побеги, или генеративные побеги могут вовсе отсутствовать. Мощные стебли диаметром 2,5 мм несут более 25–30 листьев, состоящих из 6–8 пар листочков. У особей этой онтогенетической группы, по сравнению с другими, линейные размеры годичных побегов и их органов максимальные.
Многолетняя часть куста образована остающимися из года в год базальными частями годичных побегов, образующих отдельные многолетние ветви. Максимальная длительность жизни такой ветви 6–8 лет, после ее отмирания из спящих почек, заложенных на многолетней части, вырастают новые. Отмирание ветвей начинается с центра. Основания отмерших ветвей в дальнейшем образуют партикулы, нарастание кото-
87
рых происходит по периферии. Основой куста являются партикулы, форма растения плотнокустовая с компактным каудексом. Так как отмирает большая часть побега, то истинный прирост куста очень незначителен. Вся многолетняя часть взрослого куста составляет не более 5– 10 (13) см. Длительность состояния 10–12 лет.
Для СТАРЫХ ГЕНЕРАТИВНЫХ особей характерно сокращение числа побегов в кусте, так как усиливаются процессы отмирания, уменьшается мощность побегов, снижается количество генеративных побегов до 3–8. Характерно расщепление компактного каудекса на несколько партикул, которые заметно удалены друг от друга, что приводит к преобразованию компактного каудекса в ветвистый, а куст при этом становится рыхлым. Листьев на побегах от 10 до 20 штук. Цветков в соцветии до 13 штук.
Продолжительность жизни старых генеративных особей – 6–8 (10) лет. Весь генеративный период в общей сложности составляет 7–27 лет.
Постгенеративный период характеризуется старением растения. Старые отмершие части преобладают надживыми вегетирующими побегами.
СУБСЕНИЛЬНЫЕ растения отличаются большим количеством засохших побегов. Особи в этом онтогенетическом состоянии сильно партикулируют. При подкопке особей можно наблюдать, как они распадаются на отдельные парциальные кусты, хотя внешне растение имело вид одного куста.
В настоящее время имеется три точки зрения на процесс партикуляции:
а) это форма вегетативного размножения растений (Радкевич, Шубина, 1935; Закржевский, Коровин, 1949);
б) следствие приспособления к условиям существования ( Василев-
ская, 1940);
в) признак старения и отмирания растений (Iost, 1890; Стешенко, 1960; Белостоков,1954; Казарян, Балагезян,1960; Шалыт, 1965).
Мы придерживаемся последней точки зрения и считаем, что у А. zingeri – стержнекорневого полукустарничка – партикуляция представляет собой признак старения растения, так как при образовании партикул последние отличаются слабой жизненностью и вскоре отмирают, не оставив после себя вегетативного потомства. Длительность состояния варьирует от 2–3 до 5–7 лет в зависимости от экологических условий и жизненности особей.
СЕНИЛЬНЫЕ особи представлены растениями последнего года жизни с очень ослабленной способностью развивать слабые вегетативные побеги. Партикуляция охватывает весь каудекс, что приводит к истончению партикул и постепенному разрушению каудекса. Возобновление побегов из почек на каудексе прекращается. Растение отмирает.
88
Длительность постгенеративного периода 2–4 года. Продолжительность жизни А. zingeri в среднем 15–40 лет.
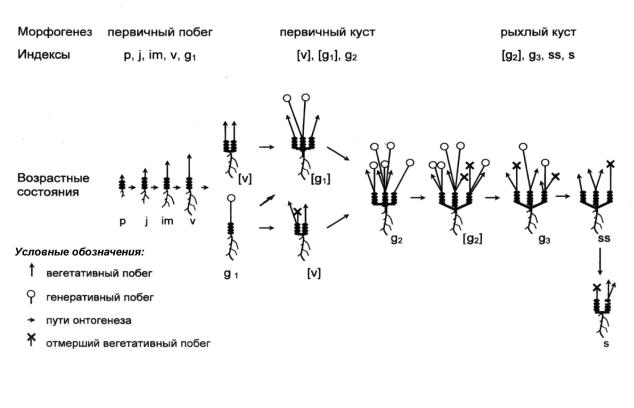
Соотношение развития жизненной формы А. zingeri с морфологическими изменениями в ходе онтогенеза позволило выделить следующие фазы морфогенеза (рис. 8):
1.Первичный побег (p, j, im, v) – от начала прорастания до отмирания верхушки моноподиального побега. Особи однопобеговые с моноподиальным типом нарастания. Тип биоморфы – моноцентрический.
2.Первичный (плотный куст) ([v], g1, g2) – с момента симподиального нарастания до начала образования рыхлокустовой структуры. Особи многопобеговые с симподиальным типом нарастания. Тип биоморфы – моноцентрический. Каудекс расположен компактно.
3.Рыхлый куст ([g2], g3, ss, s) – особи многопобеговые, частичная поздняя неспециализированная дезинтеграция. Тип биоморфы – моноцентрический. Многоглавый каудекс расположен рыхло.
Таким образом, в результате изучения онтоморфогенеза А. zingeri выяснилось, что вид относится к группе биоморф моноцентрического типа, особь в течение большей части онтогенеза существует в виде компактного единого образования и с момента поздней неспециализи-
рованной дезинтеграции наблюдается партикуляция, не приводящая к омоложению. В результате этого процесса развивающиеся побеги перемещаются на периферию. Растения вегетативно неподвижные, размножение только семенным путем (Родионова, 2003в, 2004).
Цикл фенологического развития А. zingeri складывается (рис. 9) из ряда последовательно сменяющихся ростовых и формообразовательных процессов (Родионова, 2000б).
Вегетировать растение начинает в апреле. Весной происходит быстрое развитие Аstragalus zingeri. На базальной части побега в пазухах листьев закладываются почки возобновления. Весной эти почки трогаются в рост, давая новые побеги (оси следующих порядков). Спящими остаются почки, заложившиеся в пазухах чешуй. Они обладают скрытым ростом.
К концу апреля длина побегов в природе достигает 10–18 см, на а г- робиостанции 14–24 см. В пазухах ассимилирующих листьев на уровне 4–9 междоузлия развиваются головчатые соцветия. Длина цветоноса 0,5–0,8 см. Длина самого соцветия 0,5 см.
В первой декаде мая прирост побегов резко увеличивается и в конце мая – первой неделе июня они достигают максимальных размеров. Вполне развитые побеги у средневозрастных генеративных особей имеют длину в природе 30–40 см, на агробиостанции 37–47 см и состоят более чем из 15 развитых междоузлий.
89
90
Рис. 8. Схема морфогенеза астрагала Цингера (Astragalus zingeri Korsh.)