Материал: Жукова Л.А. (отв. ред.) Онтогенетический атлас лекарственных растений. Том VII
91
Рис. 9. Схема побегообразования астрагала Цингера (Astragalus zingeri Korsh.)
Условные обозначения:  – побег;
– побег;  – бутонизация;
– бутонизация; 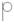 – цветущее соцвети;
– цветущее соцвети; 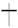 – созревание плодов;
– созревание плодов;
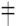 – обсеменение плодов;
– обсеменение плодов; 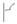 – зеленый лист;
– зеленый лист; 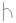 – пожелтевший лист
– пожелтевший лист
Цветение А. zingeri происходит в июне. В отдельные годы, наиболее благоприятные в климатическом отношении, единичное зацветание наблюдается в конце мая (25–30/V) и массовое цветение приходится на начало июня (7–13/VI). По характеру цветения А. zingeri является раннелетним коротковегетирующим растением. Время цветения 25–30 дней.
В кусте можно выделить следующие типы побегов:
1)почки возобновления, которые закладываются на базальных частях материнских побегов в пазухах листьев. Почки возобновления пролептического типа. Для развития полностью сформированной почки возобновления необходимо не менее 33 месяцев. Активный рост и развитие почек наблюдается в вегетационный период, предшествующий их развертыванию и совпадает обычно с переходом материнского побега к цветению. По степени сформированности побега в почках возобновления А. zingeri следует отнести к первой группе (Серебряков, 1947), то есть у него генеративная сфера сформирована к концу лета на стадии генеративных примордиев;
2)генеративные побеги в зависимости от поведения почек можно разделить на зоны: а) почки остаются спящими; б) весной следующего года трогаются в рост и дают новые однолетние побеги; в) почки трогаются в рост в этом же году, полностью реализуя себя в боковые генеративные ветви; г) почки трогаются в рост в этом же году, но боковых побегов не образуют, а формируют генеративные органы;
3)скрытогенеративные побеги – удлиненные вегетативные побеги с неполным циклом развития. У таких побегов на верхушке или в пазухах верхних листьев могут образовываться редуцированные цветки, или же верхушечная почка вытягивается, дает несколько листочков, но затем точка роста оказывается отмершей. А. П. Стешенко (1956) при изучении полукустарничков Памира пришла к выводу, что это не случайное явление. В условиях недостатка влаги оно вполне закономерно. В наиболее засушливые годы недостаток влаги и питательных веществ лишает побег возможности развиваться нормально. А. П. Стешенко назвала эти побеги переходными;
4)укороченные вегетативные побеги (брахиобласты) появляются чаще у особей с небольшим количеством генеративных побегов в авгу- сте-сентябре. В увлажненных местах число зеленых укороченных побегов выше, чем в сухих.
Как уже отмечалось, для А. zingeri характерны монокарпические побеги, которые являются моноциклическими. Наиболее мощно развиты нижние побеги, то есть у А. zingeri имеет место базитонное нарастание.
92
Во второй декаде июня образуются зеленые плоды, однако цветение особей может продолжаться. В связи с акропетальностью цветения созревание плодов и семян тоже акропетальное. Этим и объясняется наличие семян различной спелости не только в пределах популяции, но и у отдельной особи. Крайне редко можно встретить вторично цветущие особи в конце июля. В начале июля начинают созревать семена, хотя часть плодов остаются еще зелеными. Некоторые плоды поражены насекомыми, которые еще в период бутонизации проникают внутрь почки и уничтожают соцветия. Повреждают они и семена, находящиеся на самых различных стадиях развития. Растения, у которых плоды завязались и плодоношение прошло удовлетворительно, переходят к обсеменению. Этому процессу способствует влажная погода в виде кратковременного дождя с последующим быстрым переходом к высоким температурам. При высыхании плоды дружно растрескиваются. Первыми осыпаются поврежденные плоды. По способу диссеминации плодов астрагалы относятся к группе автомеханохоров. Бобы А. zingeri при созревании семян растрескиваются не полностью, а только в верхней части. Семена долго удерживаются при основании бобов, а затем опадают. Рассеивание происходит при раскачивании репродуктивных побегов, которые к периоду массового созревания плодов становятся сухими. При порывах ветра из полуоткрывшихся бобов семена рассеиваются на расстояние до 1 м. Их распространение может осуществляться и на большие расстояния при соприкосновении генеративных побегов с животными или человеком.
Вконце июля – начале августа растения полностью обсеменяются.
Вконце июля начинается процесс усыхания листовой поверхности Аstragalus zingeri, который в основном заканчивается в середине авгу-
ста. Отмирание отдельного непарноперистого листа начинается с верхушечного листочка. Полное отмирание надземной массы отмечается при сильных заморозках. Засохшие генеративные побеги долго держатся на кусте, отпадая полностью летом следующего года. Живые базальные части, несущие почки возобновления, сохраняются и ежегодно входят в истинный прирост куста. Отмирание верхней части побегов приводит к симподиальному ветвлению в кусте.
В результате исследования особенностей семенного самоподдержания А. zingeri было выявлено следующее (Родионова, 1996; 2003в):
1.Прослеживается смещение фенологических фаз развития астрагала в зависимости от географического положения популяций. Отрастание, цветение, плодоношение видов в Правобережье проходит на 5(7)– 10(14) дней раньше, чем в Левобережье.
2.По способу диссеминации вид относится к автомеханохорам.
93
3. Семена приобретают твердосемянность, что является свойством органического покоя, наступление, поддержание и выход из которого регулируются целесообразным строением кожуры.
4. В качестве предпосевной обработки семян при интродукции А. zingeri можно рекомендовать протравливание концентрированной серной кислотой от 5 до 10 минут и механическую скарификацию песком в течение 20 минут.
5.Число семяпочек в плодах является величиной относительно постоянной для вида и не зависит от условий произрастания, мало изменяется по годам.
6.Количество семян в плодах вариабельно в зависимости от фитоценотического оптимума произрастания.
7.В изученных популяциях А. zingeri слабо использует потенциальные возможности в продуцировании семян. Значительный разрыв меж-
ду ПСП и РСП объясняется высоким количеством невыполненных
инежизнеспособных семян.
8.Банк семян не является основным источником появления всходов. Основная масса проростков появляется из семян предшествующего года.
Наблюдения за фиксированными особями показали, что для А. zingeri характерен «пульсирующий онтогенез», то есть обратимые флюктуационные изменения онтогенетического состояния особи на любое другое в пределах взрослой части онтогенетического спектра. Широкое распространение «обратных» переходов в популяциях характерно
для растений, |
произрастающих в экстремальных условиях обитания |
|
(Кияк, Кобиев, |
Сварнык, 1991). Темпы развития особей |
меняются |
в зависимости |
от воздействия экзогенных и эндогенных |
факторов. |
В качестве эндогенного фактора может выступать внутрипопуляционная плотность, а в качестве экзогенного – погодные условия года. Как показали результаты наших наблюдений, при увеличении плотности особей астрагалов выше оптимальной происходит замедленное их развитие. Особи длительное время пребывают в виргинильном или временно не цветущем состоянии. Скорость прохождения онтогенетических состояний выше на склонах, чем на плакорных участках.
Относительно влияния на темпы развития погодных условий следует отметить, что 70 % особей астрагала Цингера на Малой Бахиловой горе в 1995 году вследствие жаркого лета предыдущего вегетационного сезона и сухой весны находились в состоянии вторичного покоя. В следующем вегетационном сезоне 1996 года 30 % растений скрытогенеративного состояния перешли в субсенильное, минуя средне- и старовозрастные генеративные состояния. Похожая тенденция сохранилась
94
в связи с засухой лета 2010 года в обследованной нами Каменноовражной популяции Самарского Высокого Заволжья.
Многолетние наблюдения за маркированными особями позволяют предполагать общую продолжительность онтогенеза особей вида. В караганово-разнотравно-злаковом сообществе горы Малая Бахилова полный онтогенез А. zingeri длится 15–27,5 и более лет, в разнотравноковыльном сообществе горы Стрельная – 14–24,5 и более лет. В условиях интродукции неполный онтогенез может быть пройден за 8–18 лет, при этом темпы развития особей в прегенеративном периоде значительно ускоряются, вместе с тем усиливаются процессы дезинтеграции и отмирания особей. Продолжительность пребывания А. zingeri в какомлибо онтогенетическом состоянии во многом зависит от экологофитоценотических условий его произрастания, плотности и жизненности особей (Родионова, 2001; 2003а; 2003б).
В границах изученных ценозов выявлены следующие классы динамической поливариантности темпов развития особей вида по Л. А. Жуковой (1995): нормальное, ускоренное развитие, состояние вторичного покоя, перерывов в цветении генеративных особей, омоложения и квазисенильности (Родионова, 2004). Согласно классификации типов онтогенеза цветковых растений (Жукова, 1995) онтогенез А. zingeri относится к I надтипу, А-типу, А2-подтипу: индивидуальное развитие осуществляется в течение жизни одной особи, неспособной к вегетативному размножению при наличии постгенеративного периода.
Обследованные ценопопуляции А. zingeri нормальные (гомеостатические), полночленные и неполночленные. Динамические процессы, происходящие во взрослой части популяции стабильнее, по сравнению с динамикой подроста. Наблюдается изменение индекса возрастности и численности ценопопуляций то в сторону уменьшения, то в сторону увеличения, но структура их при этом не меняется, то есть популяции находятся в устойчивом состоянии, что позволяет представить тип динамики как флюктуационный (Родионова, 2001).
95