Материал: Экспериментальное изучение нейрофизиологических механизмов процессов компенсации в случае острого повреждения стволовых структур головного мозга
Повреждение ствола мозга приводит, прежде всего, к нарушениям работы специфических систем головного мозга. Глубина нарушений функций в этом случае зависит от локализации травмы и/или степени разрушения ствола. Кроме того, степень нарушения стволовых функций обусловлена и сдвигами в системе кровообращения. В частности, очень высок риск появления сосудистой непроходимости и других нарушений церебральной гемодинамики в очень широких пределах: от ишемии до гиперемии (Фокин, 1985; Шарова, 1999). В результате повреждения ствола мозга происходит и нарушение функций стволовых регуляционных механизмов, в частности, ретикулярной формации ствола мозга, что приводит к снижению "активности" мозга (Брагина с соавт., 1988).
Экспериментальное исследование компенсаторных процессов после повреждения ствола мозга проводилось в лаборатории Э.А.Асратяна.
Так методом сечения боковой половины ствола мозга на уровне среднего и продолговатого мозга, а также используя локальное разрушение отдельных частей последнего (Незлина, 1957; Асратян, 1959), были выявлены общие закономерности восстановительных процессов в центральной нервной системе и определены их эволюционные особенности, что позволило Асратяну (1959) определить фазы развития компенсаторного процесса.
Первая фаза - травматического угнетения, возникающая в близлежащих или отдаленных по отношению к очагу повреждения структурах известная как центральный шок и выполняет, по мнению Э. А. Асратяна, охранительную и восстановительную роль, проявляя признаки запредельного торможения.
Вторая фаза (экзальтации) имеет для восстановительного процесса ключевое значение. При ней происходит усиление и генерализация специфической деятельности, а также активизация запасных центральных и проводящих нервных элементов, имеющих отношение к данной функции.
Кроме двух вышеперечисленных фаз выявлена и третья фаза: фаза умеренной возбудимости, связанная с отшлифовкой восстановленных и компенсированных функций. Е.В.Шарова (1999) считает, что "при наличии некоторых общих черт в фазах, выделяемых клиницистами и нейрофизиологами, полного тождества между ними нет, а механизмы формирования до конца не выяснены".
В работах школы Э.А. Асратяна в исследуемых восстановительных процессах ведущая роль отводилась коре. Но, сочетая удаление разных участков коры больших полушарий с повреждением ствола мозга, Э.А. Асратян отмечал решающую роль коры в восстановительных процессах при повреждении соматических функций и ее незначительную роль в процессах восстановления вегетативных функций, нарушенных в результате стволового повреждения (Асратян, 1959).
Положения Э.А. Асратяна (1959) о механизмах восстановления специфических рефлекторных стволовых нарушений уточнялись и развивались и в последующие годы (И.А.Вартанян, 1998).
По мнению Батуева (1984), от состояния неокортекса и его коррегирующих влияний во многом зависит повышение специфической и неспецифической сопротивляемости организма, выбор механизмов его активной адаптации к повреждающим факторам. Повышение устойчивости организма, как предполагается, реализуется с помощью нисходящих кортикофугальных влияний путем вовлечения в стресс-реакцию (первая фаза в посттравматический или послеоперационный период) гипоталамо-гипофизарной и гипоталамо-адреналовой системы (Макаренко, 1993). Установлена ведущая роли поля 24 передней области лимбической коры (то, что мы называем орбитофронтальной корой) в приеме и обработке висцеро-соматической афферентации, на основании существования прямых проекций нейронов лимбической коры в вегетативные структуры продолговатого мозга (Баклаваджян с соавт., 2000).
В свете вышеизложенного представляют интерес эксперименты, свидетельствующие, что активация нейронов фронтальной зоны неокортекса посредством анодной поляризации способствует повышению активности ретикулярной формации среднего мозга и других образований ЦНС, увеличивается продолжительность жизни животных в условиях передозировки различного вида наркоза (Макаренко, 1993).
Так как стволовые повреждения приводят к нарушению деятельности и активирующих систем мозга и висцеро-вегетативным сдвигам, то у больных наряду с вегетативными расстройствами могут наблюдаться психические нарушения, проявляющиеся, в частности, в повышенной эмоциональности или депрессивных состояниях (Вейн, Соловьева, 1973). Клиницисты отмечают проявление общей неспецифической реакции в раннем послеоперационном периоде. Эти реакции связаны с нарушением висцерально-вегетативных регуляций при поражении различных уровней мозга (Смирнова, 1986). При любой локализации патологического очага тяжесть клинической картины и исход заболевания зависят от вегетативных нарушений, отражающих уровень и степень вовлечения стволовых структур в патологический процесс (Полякова, 1999).
При повреждениях ствола мозга, приводящих к
изменениям функционального состояния организма, для обеспечения определенных
поведенческих актов в новых условиях (т.е. для протекания компенсаторных
процессов) необходимо участие структур лимбической системы, которые имеют
большое значение для поддержания внутренней среды организма на определенном
уровне (Ониани, 1980). Следует отметить, что при многочисленности исследований
роли неокортекса в компенсаторных процессах, участие структур лимбической
системы в компенсации нарушенных функций, в частности, при повреждении ствола
изучено недостаточно.
1.6 Значение
структур лимбической системы в формировании поведения
Как уже было сказано выше, к стволу мозга относятся структуры ЦНС, играющие ключевую роль в поддержании гомеостаза. При поражении ствола в патологический процесс вовлекаются часть интегративной системы мозга ретикулярная формация, имеющая тесные морфологические связи с гипоталамо-гипофизарной системой. Осуществление организации поведенческих реакций, направленных на приспособление к меняющимся условиям внешней и внутренней среды, возможно благодаря взаимоотношению разных частей интегративной системы. Термином «поведение» обозначается такая форма жизнедеятельности человека и животных, которая направлена на удовлетворение имеющихся у организма потребностей (Симонов, 2001). При поражении стволовых структур имеет место комплексный характер клинических нарушений, в частности, у больных вегетативные расстройства сочетаются с психическими нарушениями. Это могут быть изменения в эмоциональной сфере в виде депрессивных расстройств, либо в виде повышенной эмоциональности (Вейн, Соловьева, 1973).
Эмоция - особая форма психического отражения, которая в форме непосредственного переживания отражает не объективные явления, а субъективное к ним отношение (Данилова, Крылова, 1997). Эмоция представляет собой общее интегральное состояние организма, сопряженное с гормональными сдвигами в организме; она выражается в двигательных и вегетативных реакциях и имеет субъективную окраску в виде переживания человека его отношения к окружающему миру (Нуцубидзе, 1969). В литературе описано три уровня функционирования эмоций: 1) на уровне социальной системы эмоции облегчают поведение между различными особями; 2) подкрепление поведения на уровне отдельного организма; 3) поддержка гомеостаза на физиологическом уровне (Scott, 1980).
В нормальных условиях эмоция прочно связана с восприятием и возникает по его поводу. Основным элементами эмоций у человека являются эмоциональное состояние, которое может сопровождаться эмоциональными реакциями и вегетативными сдвигами (Вальдман, 1972а).
Существование психического компонента (эмоционального состояния) у животных допускается только по аналогии с примитивными эмоциями человека. Примитивные эмоции связаны с потребностями человека и животных в пище, в защите от повреждающих организм факторов и в размножении. Нарушение динамического стереотипа является критичным звеном для вовлечения аппарата эмоций (Симонов, 1966; Вальдман, 1972а). Так как эмоции тесно связаны с потребностями и фактически осознание потребностей, как у человека, так и у животных происходит на основании субъективных эмоциональных ощущений, то можно сказать, что потребности - это фильтр, определяющий избирательное отношение к факторам окружающий среды (Пигарева, 1983).
Эмоции делятся на положительные и отрицательные. Положительные эмоции у людей и животных описываются в процессе полового, игрового и исследовательского поведения. Отрицательные эмоции имеют место на аверсивные стимулы, на новые стимулы и в ситуации прагматической неопределенности (Azrin et al., 1965; Cahoon, 1972; Karczmar et al., 1978). В этих случаях имеет место реакция страха. Страх мотивирует выработку реакции избегания, в чем заключается его адаптационное значение. Реакция страха может выражаться как в агрессии, так и в двигательном торможении. В литературе имеются данные, что агрессивные реакции стимулируются новизной стимула, которые как увеличивают агрессивность, так и направляют ее. Проявление агрессивного поведения увеличивает общую активность организма. По мнению Пигаревой (1983) - одна из функций эмоций -подкрепление поведения, являющегося адаптивным в данных условиях. Одно и то же физиологическое состояние может давать разные эмоциональные проявления в зависимости от окружающих условий и индивидуальных особенностей. Кроме того, поведенческое реагирование обусловлено еще и доминирующей мотивацией. Но кроме внутренних стимулов реализация поведения зависит от адекватных внешних условий (Пошивалов, 1986). Таким образом, в поведенческие акты животных входят эмоциональные реакции, которые определенным образом способствуют целесообразному приспособлению к условиям среды (Вальдман, 1972а).
Изучая отражательную функцию эмоций и придавая решающее значение оценке вероятности удовлетворения потребности в генезе эмоций, П.В. Симонов в 1964 году создал «потребностно - информационную теорию эмоций», согласно которой эмоции высших животных и человека определяются какой-либо актуальной потребностью и оценкой вероятности удовлетворения этой потребности (Симонов, 2001). Отрицательные эмоции, по П. В. Симонову, возникают, когда мотивированное поведение сталкивается с препятствием. Устранение препятствия или его преодоление сопровождается положительными эмоциями (Симонов, 1993).
Генерирование эмоций в ситуации прагматической неопределенности имеет адаптивное значение. При возникновении эмоционального напряжения развиваются вегетативные сдвиги, объем которых, как правило, превышает реальные потребности организма. Таким образом, биологическое значение эмоций заключается в подготовке организма для активности (избыточная мобилизация энергетических ресурсов) и в замещении недостатка информации. В этом и заключается компенсаторная функция эмоций (Нуцубидзе, 1969; Симонов, 2001).
Вопрос о «субстрате эмоций» определенно может быть решен только в отношении топографии интегративных центров (зон) эмоционально-выразительных проявлений (Вальдман, 1972).
Нейроанатомия эмоций.
Эмоция, - сложное образование, предполагающее интеграцию соматических, вегетативных и субъективных компонентов (Данилова, Крылова, 2000). Первая наиболее стройная концепция, связывающая эмоции с функциями определенных структур мозга (т.н. круг Пейпеца), была опубликована в 1937 г. Дж.Пейпецом. Изучая эмоциональные расстройства у больных с поражением гиппокампа и поясной извилины (Пейпец, 1962), автор выдвинул гипотезу о существовании единой системы, объединяющей ряд структур мозга и образующей мозговой субстрат для эмоций. В дальнейшем были описаны нарушения эмоциональных реакций у больных при повреждении медиальных отделов височной доли (Доброхотова, 1974; Мадорский, 1985)
Наиболее полная схема взаимодействия мозговых
структур в процессе организации поведенческого акта разработана П.В.Симоновым
(рис. 1.1).
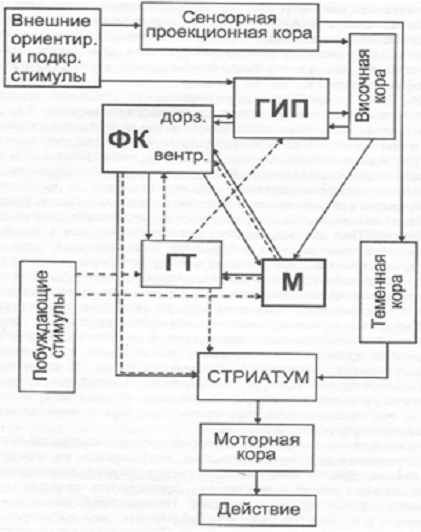
Рис. 1.1 Схема взаимодействия мозговых структур в процессе организации поведенческого акта.
Условные обозначения: ФК - фронтальная кора; ГИП - гиппокамп; М -миндалина; ГТ - гипоталамус.
Сплошные линии - информационная афферентация; прерывистые -мотивационные влияния; двойные - эмоционально-окрашенная афферентация. (П.В. Симонов, 2001)
По мнению автора, все многообразие эмоциональных состояний реализуется конкретным набором структур мозга, отнесенным к «информационному» и «мотивационному» блокам (Симонов, 1993, 2001).
Из всех структур круга Пейпеца наиболее тесную связь с эмоциональным поведением обнаруживают гипоталамус (мотивационная структура по П.В. Симонову) и поясная извилина, относящаяся к информационному блоку. Кроме того, оказалось, что и другие структуры мозга, не входящие в состав круга Пейпеца, влияют на эмоциональное поведение. Среди них особая роль принадлежит миндалине (мотивационный блок), а также лобной и височной коре головного мозга.
В составе эмоции принято выделять собственно эмоциональное переживание и его соматическое и висцеральное выражение; возможность их появления независимо друг от друга свидетельствует об относительной самостоятельности их механизмов. Так, эмоциональное выражение весьма устойчиво к действию транквилизаторов, в отличие от эмоционального переживания (состояния), которое ими легко угнетается (Вальдман с соавт., 1976). Диссоциация эмоционального переживания и его выражения в двигательных и вегетативных реакциях обнаружена при некоторых поражениях ствола мозга. Она выражается в так называемых псевдоэффектах: в этом случае интенсивные мимические и вегетативные реакции, характерные для плача или смеха, могут протекать без соответствующих субъективных ощущений (Вальдман с соавт., 1976).
В регуляции эмоций особое значение имеют корковые структуры. Повреждение поясной извилины в эксперименте оказалось наиболее эффективным в отношении появления эмоциональных реакций. Так у животных с поврежденной передней частью поясной извилины имел место феномен «потери страха» (Glees et al., 1950); в случае болевого раздражения реакция страха заменялась реакцией гнева (Ониани, 1980). Наиболее яркие поведенческие реакции оборонительного типа наблюдались при повреждении передней части поясной извилины (Ониани, 1980; Cardinal et al., 2002). У человека поражение лобных долей приводит к глубоким нарушениям эмоциональной сферы (Gainotti, 1983; Jorge, Robinson, 2003; Jorge et al. 2004). Предполагается, что фронтальные отделы коры осуществляют свое модулирующее влияние на эмоциональные реакции благодаря связям с миндалевидным комплексом и гипоталамусом (Wolf, Sutin, 1966; Моторина, 1968; Nauta, 1972; Schoenbaum et al., 1999). Согласно современным данным, поясная извилина имеет двусторонние связи со многими подкорковыми структурами (перегородкой, верхними буграми четверохолмия, голубым пятном и др.), а также с различными областями коры в лобных, теменных и височных отделах (Ward, 1948; Замбржицкий, 1972; Nauta, 1972; Maurice et al., 1999). Некоторые авторы высказывают предположение о высшей координирующей функции поясной извилины в отношении эмоций (Ониани, 1980; Баклаваджян с соавт., 2000).
В настоящее время накоплено большое число экспериментальных и клинических данных о роли полушарий головного мозга в регуляции эмоций. Изучение функций левого и правого полушария выявило существование эмоциональной асимметрии мозга. Так, Брагина и Доброхотова (1981) установили, что больные с поражениями в левом полушарии тревожны, озабочены, тогда как правостороннее поражение сочетается с легкомыслием, беспечностью.
К информационному блоку относится также гиппокамп, в котором, по мнению П.В. Симонова (1993) происходит обработка внешних сигналов, требующих эмоционального реагирования, сопоставление их с опытом, хранящимся в памяти, и определение стратегии необходимых действий.
Однако данные по изучению влияния повреждения гиппокампа на мотивационно-эмоциональное поведение животных менее отчетливы, чем таковые при повреждении поясной извилины (Ониани, 1980).
Так, О. С. Виноградова (1975) возражает против участия гиппокампа в механизмах эмоциональных реакций, основываясь на противоречивости экспериментальных и клинических данных. Сложность трактовки результатов, полученных методом локального повреждения мозга, обусловлено неизбежным внедрением в систему анатомически и функционально связанных структур мозга (Пигарева, 1984; Abe, 2001). Следует отметить существование большого количества работ, свидетельствующих о тормозном влиянии гиппокампа на возбудимость различных структур мозга (Roberts et al., 1962; Зилов, Рогачева, 1975; Лагутина и др., 1976).
С другой стороны, существуют исследования, в
которых демонстрировалось появление эмоциональных реакций после повреждения
гиппокампа (Гамбарян, Коваль, 1973), корреляция гиппокампального тета-ритма с
мотивационно-эмоциональными реакциями (Хананашвили, 1972). По данным Умрюхина
(2002), гиппокамп принадлежит к числу надгипоталамических структур,
регулирующих активность гипоталамо-гипофизарно-надпочечниковой системы, что
заставляет еще и еще раз возвращаться к проблеме функций гиппокампа. Эффект
воздействия на гиппокамп зависит как от функционального состояния организма,
так и от условий внешней среды (Пигарева, 1978; 1984), поэтому важно
разобраться в обилии противоречивых фактов при исследовании роли этой структуры
в интегративной деятельности мозга при адаптивно-компенсаторных перестройках в
случае локального стволового повреждения.
Глава 2. Объект и методы исследования
Настоящая работа посвящена изучению динамических системных компенсаторных реакций мозга крысы, развивающихся в ответ на локальное повреждение ствола мозга. Решение этой задачи проводилось в следующих направлениях:
1. Создание адекватных экспериментальных физиологических моделей острого повреждения ствола мозга крысы.
2. Динамический анализ неврологических нарушений и поведенческих реакций при изолированном и сочетанном стволовом повреждении.
3. Электрофизиологический анализ системных нейродинамических перестроек ЦНС в ответ на локальное стволовое повреждение.