Материал: Экспериментальное изучение нейрофизиологических механизмов процессов компенсации в случае острого повреждения стволовых структур головного мозга
Все исследования выполнены на 117 (табл. 2.1)
беспородных белых крысах-самцах и на 10 крысах линии Wistar
весом 200-300 гр.
Таблица 2.1. Сводная таблица распределение
беспородных крыс по группам
Изолированный
ствол (в т.ч. с регистрацией ЭЭГ)
Ствол
+ ОФК
Ствол
+ гиппокамп
Двустороннее
справа
слева
Двустороннее
справа
слева
28+8
16
11
10
25
9
10
Остановимся подробнее на методических аспектах
каждого из направлений исследования.
.1 Моделирование
острого локального и сочетанного стволового повреждения
Работа состояла из нескольких этапов:
). Создание экспериментальной модели
изолированного повреждения ствола мозга крыс с воспроизводимыми позными и
двигательными нарушениями, сопоставимыми с неврологической симптоматикой
очагового поражения ствола мозга человека - для последующего длительного
динамического наблюдения за состоянием животных. При этом мы руководствовались,
в частности, данными Р. Магнуса (1962) о том, что перерезка ствола на уровне
ядра Дейтерса сопровождается позными нарушениями, которые сходны с
неврологическими симптомами у больных со стволовой патологией. Для создания
экспериментальной модели проводили электролитическое разрушение ствола мозга
крыс на уровне латерального вестибулярного ядра Дейтерса (ВЯД). Сложность
выполнения этой работы определяется близостью IV-го
желудочка с расположенными там дыхательным и прессорным центрами (Thompson,
1972; Вальдман, Цирлин, 1981), нарушение целостности которых вызывает
высоковероятную гибель животных.
Сходная модель была использована Г.Н.
Крыжановским (1980) при создании генератора патологического возбуждения в
системе вестибулярных ядер. В ВЯД вводили столбнячный токсин - с последующим
электролитическим разрушением этой зоны постоянным током 5 мА длительностью
10-80 сек, что приводило к гибели животного.
В наших исследованиях подбиралась такая сила
тока, при которой повреждение ствола не влекло за собой немедленную гибель
животных. 24 крысам, находящимся под нембуталовым наркозом (50 мг/кг
внутрибрюшинно), в ствол мозга слева вводили стальной электрод диаметром 0,2 мм
по координатам Р = -10.5; L
= 3; H = 7 (согласно
атласу Pavlovits,
Bronstein,
1988) и в течение 20 секунд пропускали постоянный ток силой 1,5 мА. Это
приводило к локальному повреждению ствола на уровне латерального вестибулярного
ядра Дейтерса (рис.2.1.В). Далее мы проводили детальный анализ последствий
операции с изучением развивающихся в ЦНС адаптивно-компенсаторных реакций.
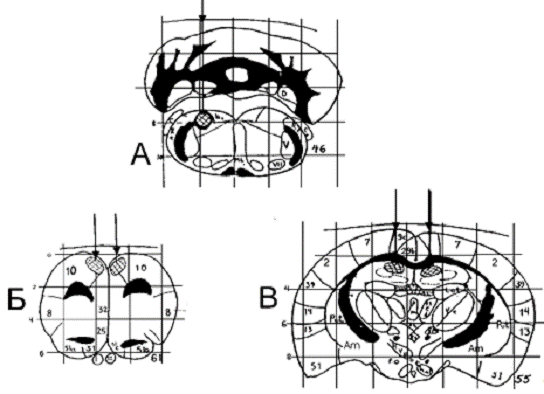
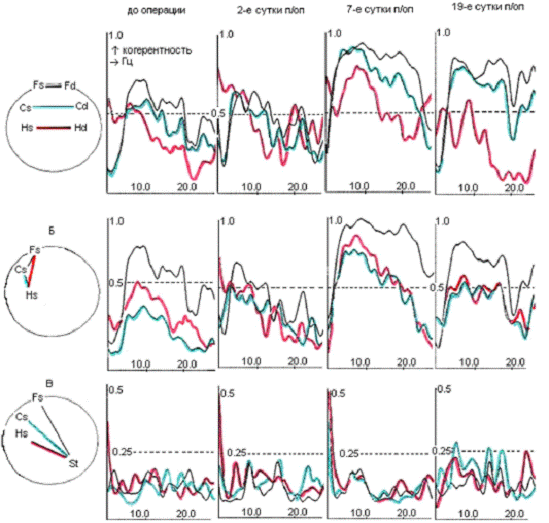
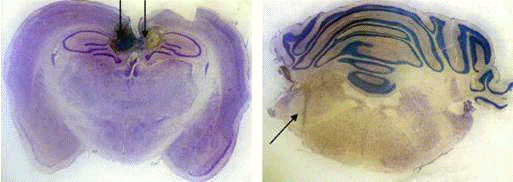
Рис.2.1 Схема стволового (А), а также
сочетанного стволово-орбитофронтального (А, Б) и стволово-гиппокампального (А,
В) разрушения мозга крыс.
Топография из атласа W.Krieg,
1946.
На схемах: цифрами отмечены номера полей,
стрелками - ход коагулирующих электродов; заштрихованные участки - зона
планируемого разрушения.
). Для выяснения роли орбитофронтальной коры
(ОФК) и гиппокампа (Гип) в компенсации функций при повреждении ствола были
разработаны модели сочетанного повреждения ствола и исследуемых структур
(рис.2.1).
Проводилось одностороннее электролитическое
повреждение ствола на уровне ядра Дейтерса (как модели послеоперационной
стволовой патологии), в сочетании с билатеральным повреждением (рис.2.1А, В)
поля СА1 гиппокампа (15 животных) или (рис.2.1А, Б) орбитофронтальной коры (16
животных).
Выбор координат повреждения гиппокампа (P=3;
L=1; H=3)
обусловлен тем, что волокна этого отдела гиппокампа входят в систему дорсального
свода, считающегося основным образованием, связывающим гиппокамп с
мезодиэнцефальными структурами (Ониани, 1980). По-видимому, это обеспечивает
ему роль модулятора эмоциональных, висцеро-вегетативных и биоэлектрических
реакций (Болдырева с соавт., 1972; Хамильтон, 1984; Симонов 1993). Медиальный
участок поля СА1 гиппокампа повреждался силой тока 0.5 мА и длительностью 20
сек.
Повреждаемые отделы ОФК (A=3;
L=0.5) соответствуют
префронтальной коре обезьян, которая определяется как проекционная зона медиодорсального
ядра таламуса (Пигарева, 1984; Ongur,
Price, 2000). Это
является аналогом средних отделов поля 24 и рассматривается рядом авторов как
лимбическая кора (Василевская, 1971; Divac,
1972; Замбржицкий, 1972; Беллер, 1977). Коагуляция ОФК осуществлялась силой
тока 0.5 мА, действующего в течение 20 секунд.
При сочетании разрушения ствола с двусторонним
повреждением исследуемых структур лимбической системы методика подвергалась
определенной модификации. У группы из 7 животных билатеральное разрушение орбитофронтальной
коры или гиппокампа следовало через 6 часов или через сутки после
одностороннего повреждения ствола мозга. В дальнейшем для уменьшения
вероятности гибели животных от суммарного объема одновременно поврежденного
мозгового вещества (Thompson,
1972) у 21-й крысы второе разрушение отставлялось на 7 дней относительно
первого. При этом изменяли и последовательность повреждения структур. То есть
сначала производили коагуляцию исследуемых структур лимбической системы, а
затем - ствола мозга.
Нами исследовались также особенности участия
право- и левосторонних образований ОФК и Гип в восстановительных процессах при
повреждении ствола. Для этого у 19 животных за 7 суток до электролитического
повреждения ствола проводилась односторонняя (справа или слева) коагуляция ОФК
током 0,5 мА, длительность коагуляции 20 сек по координатам, указанным выше. У
20 крыс в те же сроки и теми же параметрами тока производилась одностороннее
повреждение поля СА1 гиппокампа.
.2
Анализ функционального состояния животных до и после изолированного и
сочетанного стволового повреждения
У всех оперированных животных проводилась
динамическая клинико-поведенческая оценка функционального состояния в
сопоставлении с дооперационным статусом.
Перед операцией все животные взвешивались, их
эмоциональное состояние оценивали по шкале Кинга (King,
1958), определяли предпочитаемую конечность, исследовали направление движения и
активность в открытом поле (Айрапетянц с соавт., 1980; Микляева, 1989; Буреш с
соавт., 1991). При исследовании эмоциональных реакций по шестибальной шкале
оценивали формализованные качественные показатели реакции на касание пинцетом
спины и носа, взятие в руки (в частности, величину мышечного сопротивления),
уровень вокализации. Кроме того, по числу болюсов и уринаций во время
тестирования животного определяли величину эмоционального возбуждения (Левшина
с соавт., 1997; Sullivan,
Gratton, 1999).
Для выявления типологических особенностей высшей
нервной деятельности экспериментальных крыс использовали тест открытого поля.
Площадка диаметром 1 м с высотой бортов 30 см
была поделена на 36 квадратов. В центральной части были прочерчены еще две
окружности с радиусами 25 и 40 см. Центры открытого поля и этих окружностей
совпадали. Животных помещали внутрь площадки у одного из бортов. При этом
голова крысы была направлена в центр поля. Общее время наблюдения три минуты.
За это время регистрировали количество пересеченных квадратов, что служило
характеристикой уровня горизонтальной активности. Проводилось ранжирование
уровня горизонтальной активности: количество пересечений больше 50 - высокий
уровень, от 50 до 30 - средний и ниже 30 - низкий уровень горизонтальной
активности. Кроме того, регистрировали количество выходов за первый и второй
круг, длительность и частоту груминговых реакций, количество принюхиваний, а
также количество вертикальных стоек (вертикальная активность). Вертикальная
активность была низкой, если количество вставания на задние лапы было меньше
10, средней - от 10 до 20 и высокой, если количество стоек было больше 20.
Определяли также наличие и характер моторной
межполушарной асимметрии. Для этого выявляли предпочитаемую конечность и
предпочитаемое направление движения крыс в открытом поле. Направление движения
определяли по отношению общего количества пересеченных квадратов к количеству
квадратов, пересеченных при движении в одном направлении.
Предпочитаемую конечность устанавливали по
методике, описанной в работе Микляевой (1989) с небольшими изменениями,
относящимися ко времени пищевой депривации. Животных, содержавшихся на
ограниченном пищевом рационе в течение 24 часов, помещали в плексигласовую
камеру с металлической крышкой в виде сетки. На торцевом конце камеры, на
уровне 3 см от пола находилось отверстие с трубкой для подачи корма (рис.2.2).
Рис.2.2 Камера для определения предпочитаемой
конечности
Приманка подавалась с помощью инъекционной иглы
длиной 15 см. Корм в трубке располагался на расстоянии 1 см от края отверстия
таким образом, чтобы животное могло его взять только лапой.
Для исключения элемента обучения опыты
проводились в течение двух экспериментальных дней - по три предъявления пищи в
каждый из них. Животных, которые брали приманку только левой лапой,
квалифицировали как левшей. Если корм брался только правой лапой, то эти крысы
рассматривались как правши. Амбидекстрами считали животных, у которых хотя бы в
одной из шести проб менялось предпочтение используемой конечности (Микляева,
1989).
После операции проводили ежедневное наблюдение
за общим
состоянием
животных, отмечая изменения в весе, характере двигательной
активности
и эмоциональной сфере, состоянии кожного покрова и шерсти.
Анализировали также послеоперационные изменения
неврологического статуса: появление
позных, мышечных и двигательных нарушений, а также
степень
и скорость восстановления нарушенных функций. Из-за тяжести
состояния
после операции коагуляции ствола мозга повторное тестирование в
открытом
поле было возможно не у всех особей.
Сопоставление состояния и поведения животных до
и после повреждения головного мозга со статистическим анализом использованных
признаков проводили совместно с к.б.н. М.А.Куликовым. Использовались методы
статистической обработки из пакета программ STATISTICA.
В основном - методы рангового (порядкового) анализа - для сравнения двух
независимых групп критерии Манна-Уитни или Вилкоксона, для сравнения долей -
критерий хи-квадрат, для сравнения нескольких групп -непараметрический
дисперсионный анализ Крускалла-Уэллиса, для оценки связи двух показателей -
критерий ранговой корреляции Спирмана. Параллельно с этим для наглядности и
контроля результатов использовались и аналогичные параметрические критерии -
Стьюдента, Пирсона, однофакторный дисперсионный анализ (ANOVA)
с последующей проверкой различия средних по критерию Стьюдента-Ньюмена-Келлса.
В подавляющем большинстве случаев статистические выводы при применении этих
двух групп методов совпадали.
2.3
Нейрофизиологический анализ адаптивно-компенсаторных реакций ЦНС при локальном
стволовом повреждении
У 8 животных в 27 опытах были проведены
динамические электрофизиологические исследования до и после одностороннего
электролитического повреждения латерального вестибулярного ядра Дейтерса. Они
включали одновременную регистрацию электрической активности (ЭА) коры, гиппокампа,
а также интактного ВЯД - с ее последующим спектрально-когерентным анализом.
(Исследования выполнены совместно с д.б.н. Е.В.Шаровой).
Регистрацию ЭА проводили платиновыми электродами
диаметром 0,2 мм, напаянными на штырьки. Место припоя и электрод по всей длине
(за исключением кончика) изолировались тремя слоями винифлексового лака.
Регистрирующие электроды вживляли в симметричные
лобные (А=3; L=0,5),
соматосенсорные (А=2; L=3,5)
области коры, симметричные области поля СА1 гиппокампа (Р=3; L=1;
H=3) и интактное
латеральное вестибулярное ядро Дейтерса (Р=10,5; L=3;
H=7).
Запись биопотенциалов производили в
экранированной установке. В плексигласовую камеру с заземленным металлическим
полом помещали животное, предварительно адаптированное к обстановке. ЭА
регистрировали на энцефалографе фирмы "Медикор" (Венгрия)
монополярно. Индифферентный электрод, вживлялся в носовую кость.
Регистрировался сигнал в диапазоне 0,5 -30 Гц. Запись осуществляли при
состоянии спокойного бодрствования животного с открытыми глазами. Кроме того,
исследовали реакции крыс на значимые для данного вида животных слуховые
раздражители (шелест бумаги и громкий голос человека) до операции (запись фона)
и после повреждения ствола на уровне ВЯД. Исследование ЭА проводили по
следующей схеме. После вживления электродов ЭА записывали 2-3 раза в течение
недели. На данном этапе эксперимента животные могут рассматриваться как
ложнооперированные. Убедившись в отсутствии у крысы поведенческих и
неврологических нарушений, а также в стабильности паттерна фоновой ЭА,
осуществляли стволовую коагуляцию. После операции ЭА регистрировали в динамике:
на 1-3, 5-7, 9-12-е сутки и далее с увеличивающимися интервалами. Длительность
наблюдения (от первых суток до месяца) и число индивидуальных исследований (от 2
до 5) определялась как особенностями функционального состояния животного, так и
плотностью примыкания колодки к кости черепа, которая уменьшалась при
подрастании соединительной и мышечной ткани.
Одновременно с регистрацией ЭА на энцефалографе
проводили запись ее на жестком диске персонального компьютера в программе
"Нейрокартограф" отечественной фирмы МБН - для их последующего
спектрального и когерентного анализа. Трехминутные реализации ЭА обрабатывали
на вычислительном комплексе фирмы МБН (Россия). При этом спектры когерентности
рассчитывались для симметричных областей полушарий и гиппокампа, для отведений
внутри каждого полушария, а также для интактной стволовой области с упомянутыми
выше отделами обоих полушарий. Рассматривали также фазовые спектры анализируемых
пар отведений.
Методической особенностью нашего
электрофизиологического исследования являлся раздельный качественный и
количественный анализ двух паттернов, характерных для ЭА мозга крысы:
синхронной и десинхронной составляющей. По мнению Буреша с соавторами (1991),
эти две составляющие, отражают разное функциональное состояние животных:
дремоту и бодрствование соответственно. Длительность отредактированных и
готовых к математической обработке фрагментов ЭА составляла не менее одной
минуты.
Спектры мощности синхронной и десинхронной
составляющих ЭА вычисляли на основе быстрого преобразование Фурье. При
спектральном анализе определялась энергия каждой частотной составляющей с
разбивкой их по диапазонам физиологических ритмов (дельта, тета, альфа и бета).
После обработки отрезков ЭА на выходе получали графики спектральной мощности, в
которых на оси абсцисс отложены значения частоты биоэлектрической активности в
Гц, по оси ординат - ее мощности в мкВ , вычисляемая для данной частоты за
анализируемый отрезок времени.
Параметры и вид энергетических спектров
позволяют судить о выраженности отдельных ритмов в определенных структурах
мозга при разных функциональных состояниях, распределении ЭА внутри и между
полушариями.
Кроме того, определяли уровень согласованности
и, следовательно, взаимосвязанности определенных зон мозга, показателем чего
является функция когерентности. Показано, что определенный (оптимальный)
уровень сочетанности биопотенциалов является нейрофизиологическим базисом для
реализации специфических функций ЦНС (Гриндель, 1980; Русинов с соавт., 1987;
Ливанов, Думенко, 1987; Ливанов, 1989). Для количественной оценки использовали
значения когерентности отдельных частотных диапазонов, а также средний уровень
когерентности диапазона 0,5 - 20гц (Русинов с соавт., 1975). Величина
когерентности равна 0 для полностью независимых процессов и 1 при их абсолютной
идентичности, т.е. совпадению по частоте при одномоментности появления
(Гриндель с соавт., 1973).
Временные отношения отдельных ритмов - опережение
или отставание каждого ритма в двух отделах головного мозга, определяется
посредством анализа фазового спектра. По мнению ряда авторов (Гриндель с
соавт., 1973; Ливанов, 1989), сдвиг фаз характеризует разность
последовательности вовлечения структур в интегративную деятельность. На
графиках фазовых спектров выявлялись опережающие зоны регистрации по частотам
относительно нулевой линии. При фазовом сдвиге вверх относительно нулевой линии
"ведущим" являлось первое из пары отведений. Достоверность изменения спектрально-когерентных
характеристик электрической активности до и после операции оценивалась по
оригинальной программе В.Г. Воронова,
О.М.
Гриндель
и И.Г.
Скорятиной,
которая основана на Т-критерии Стьюдента (Воронов, 2000, c.244-245).
Кроме регистрации биоэлектрической активности,
для объективной оценки функционального состояния у части животных проводились
исследования работы сердца до и после повреждения ствола. Для этого у 3 крыс
регистрировалась частота сердечных сокращений (ЧСС) по модифицированной методике
Н.Г.
Михайловой
(Михайлова, 1994). Регистрация производилась с помощью эластичного электрода в
хлорвиниловой оболочке, со свободным от изоляции серебряным шариком на конце,
вживленным под кожу в верхней части спины. С другой стороны электрод заканчивался
штырьком, зафиксированным пластмассой на черепе животного. Регистрация ЧСС
производилась на энцефалографе с параллельной регистрацией на жестком диске
персонального компьютера. Обработка полученных данных производилась по
программам спектрального анализа параллельно с обработкой записей
биопотенциалов.
2.4
Морфологические и патоморфологические исследования мозга
крыс
По истечении двух недель или полутора месяцев
выживших животных с односторонней коагуляцией ствола мозга и/или через два -
два с половиной месяца с сочетанными разрушениями усыпляли парами эфира,
декапитировали; мозг фиксировали в 10 процентном растворе формалина. В
формалине мозг выдерживался не менее недели, после чего изготовляли серийные
срезы толщиной 30 мкм и окрашивали их по Нисслю. На препаратах определяли
локализацию и размер разрушений и сопоставляли результаты морфологического
анализа с данными поведенческих и электрофизиологических исследований. Работа
проводилась совместно с Т.С.Михеевой и В.Н.Мац.
В случае гибели животного, помимо
морфологических, проводились патологоанатомические исследования
макроскопической оценки состояния оболочек мозга, самого нервного субстрата и
состояния внутренних органов для установления причины гибели животных. Затем
мозг заливался в целлоидин, срезы окрашивали гематоксилином и эозином и
анализировали для выявления причин
смерти и возможных расстройств кровообращения. Этот этап работы проводился на
базе Института нейрохирургии им. Бурденко ст.н.сотр. этого Института Шишкиной
Л.В.
2.5 Эффекты изолированного
стволового повреждения мозга крыс
Данный раздел работы посвящен анализу системных
реакций ЦНС, развивающихся в ответ на острое локальное повреждение ствола мозга
крыс, имеющих, в целом, компенсаторную направленность.
Несмотря на многочисленные как клинические
(Благовещенская 1957; Майорчик, 1973; Дубикайтис, Полякова, 1983; Rath,
Klein, 1991), так и
экспериментальные исследования (Незлина, 1957; Асратян, 1959; Подачин с соавт.,
1983), адаптивно-компенсаторные механизмы, лежащие в основе восстановительных
процессов мозга при остром повреждении его ствола, изучены недостаточно. Вместе
с тем, их анализ сохраняет теоретическую и практическую значимость - в
частности, в связи с большим числом больных, перенесших такое повреждение
(сосудистого, травматического и др. генеза) и нуждающихся в реабилитации.
В нашей работе мы изучали внешние
(неврологические, соматические, поведенческие), а также электрографические
проявления церебральных реакций животных после острого поражения ствола
головного мозга. Проведение экспериментальных исследований по этой проблематике
представляется нам важным и целесообразным, т.к. они позволяют ответить на ряд
вопросов, возникающих, но неразрешимых (по этическим и методологическим
причинам) в клинических исследованиях на человеке.
Основной экспериментальный материал раздела
получен на 28 животных с локальным электролитическим повреждением ствола на
уровне латерального вестибулярного ядра Дейтерса (см. главу 2).
Так как задачи нашей работы в значительной
степени определялись проблематикой нейрохирургической клиники человека (Шарова
с соавт., 1991, 1993), важным представлялось сопоставление внешних последствий
локального стволового повреждения у животных с клинической картиной у больных с
острой и хронической стволовой патологией.
Глава 3. Клинико-поведенческие последствия
локального электролитического стволового повреждения
3.1 Локальное
электролитическое повреждение ствола мозга крыс как модель экспериментальной
стволовой патологии
Изолированная электролитическая коагуляция ствола
проведена у 28 животных. По характеру послеоперационного течения и исходам все
они были разделены на три группы (таблица 3.1).
Таблица 3.1 (2) Распределение животных с
изолированным стволовым повреждением по
исходам.
Исход
Неосложненный
Осложненный
Летальный
Количество
Животных
А
Б
На операционном
столе
Всего!
28
7
5
3
4
12
(43%)
9
(32%)
7
(25%)
Первую группу составляли 12 крыс, у которых
изменения в позе и характере двигательной активности при отсутствии специальных
тестов вообще не выявлялись (подгруппа А - 7 животных), либо имели быстро
обратимый характер (подгруппа Б). Наиболее характерным неврологическим
нарушением, отмечаемым у 4 крыс подгруппы Б, было изменение позы в форме
незначительного наклона головы (не более 30 градусов) в ту сторону, на
которой
была проведена операция (рис. 3.1. А). У таких животных нормальное положение
головы восстанавливалось в течение 1-3 суток (рис. 3.1.Б). У одного животного
при этом наблюдалось также нарушение движений в виде возможности перемещаться
лишь в одну, ипсилатеральную операции (левую), сторону. Это нарушение также
регрессировало через трое суток после операции. Послеоперационное течение у
животных первой группы было расценено как неосложненное.
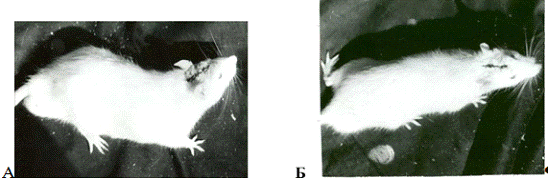
Рис.3.1. Крыса №30 с неосложненным течением
послеоперационного периода Ко второй группе (с осложненным течением) были
отнесены 9 крыс, у которых локальное
электролитическое повреждение ствола сопровождалось
стойкими
изменениями позы и двигательной активности. Нормализация неврологического
статуса была более длительной (до 13 суток) по сравнению с первой группой, а
нарушения были более тяжелыми.
Только у двух крыс из 9 послеоперационная
симптоматика сводилась к незначительному (до
30 градусов) наклону головы, который, однако, регрессировал только к 13 суткам.
У двух особей угол наклона был более выраженным
(до 90 градусов). Одновременно у пяти животных данной группы имели место более
тяжелые характерные нарушения движения в виде винтообразного перемещения влево
(рис. 3.2), которые могли развиваться не сразу. Эти винтообразные или
бочкообразные
вращения (barrel
rotation) сходны с
описанными M.L.Chon
& M.Chon
(1975) вращениями вдоль продольной оси тела, возникающими после введения
соматостатина. Сходные явления наблюдал Г.Н.Крыжановский (1980) при создании
генератора патологического возбуждения в системе вестибулярных ядер (см. главу
2). В двух наблюдениях данной группы отмечены колебания в степени выраженности
описанных нарушений: к 4 и 10 суткам после операции винтообразные движения
исчезали. В двух самых тяжелых случаях, в период с первых до четвертых суток,
животные не могли самостоятельно, не цепляясь за прутья, удерживаться на
гладкой поверхности; они лежали на полу и при потере опоры начинали вертеться
вокруг своей оси. У одного животного неврологический дефект сопровождался также
асимметричным
повышением тонуса левых конечностей.
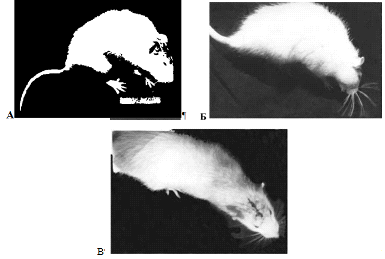
Рис 3.2 Крыс № 26 с осложненным течением
послеоперационного периода А - третьи
сутки после операции; Б - шестые сутки после операции; В -четырнадцатые сутки
после операции
Описываемые неврологические нарушения
проявлялись волнообразно: кратковременное улучшение могло чередоваться с
нарастанием симптоматики (рис. 3.3).
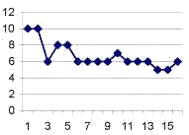
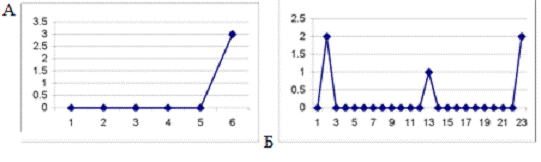
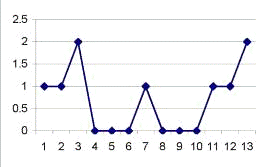
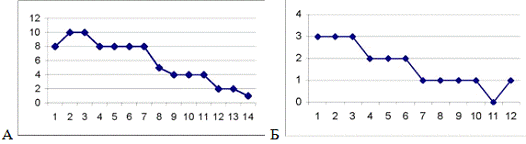
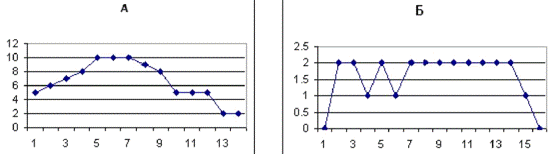
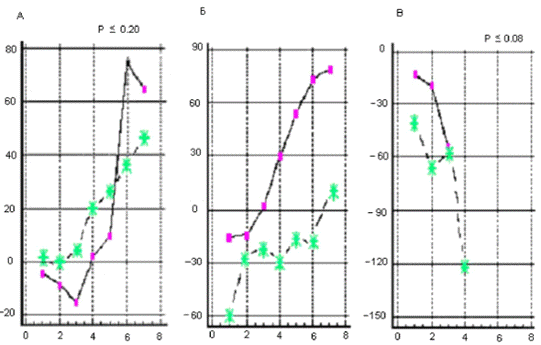
Рис. 3.3. График, отражающий характер течения
осложненного послеоперационного периода у крысы №18. По оси абсцисс - сутки
после операции. По оси ординат - ранжированные показатели неврологических
отклонений. Максимальные отклонения в виде винтообразных движений соответствуют
10 баллам.
Еще в четырех наблюдениях неврологический дефект
не компенсировался полностью до усыпления животного. Указанные обстоятельства и
позволили нам расценить послеоперационное течение у животных второй группы как
осложненное.
В третью группу были включены семь крыс, для
которых локальное электролитическое повреждение ствола закончилось летальным
исходом. Они погибли либо на операционном столе (4 крысы), либо в разные сроки
после операции (на 4, 6 и 14-е сутки).
В трех последних случаях у животных после
повреждения ствола мозга выявлялись единообразные неврологические нарушения:
наклон головы относительно туловища под углом 90 градусов в оперированную
сторону; вытягивание правых задних конечностей; при попытке поступательного
перемещения - "винтообразное" движение. Вокруг глаз появлялись
сукровичные подтеки. Характер послеоперационного течения, как и в предыдущей
группе, был волнообразным - без видимого улучшения состояния
(рис.
3.4- фотография и график) либо с проградиентным нарастанием тяжести к моменту
гибели.
Рис. 3.4 График течения послеоперационного
периода у крысы №11, умершей на 13-е сутки после операции. Обозначения такие
же, как на рис.3.3.
Следует отметить, что ни в одной из выделенных
групп не было выявлено значимой потери веса животных в послеоперационном
периоде. Хотя незначительное его снижение (на 10-20 граммов) в первые сутки
после операции с последующим восстановлением имело место практически у всех
крыс и является, по-видимому, частью неспецифической реакции организма на
повреждение мозга.
3.2 Результаты
морфологического контроля
Сопоставление внешних (неврологических и
двигательных) эффектов повреждения с результатами морфологического контроля
свидетельствуют об определенном соответствии между локализацией и степенью
электролитического разрушения мозга, а также характером послеоперационных
нарушений.
Так, морфологические исследования не выявили
точного попадания коагулирующего электрода в левое вестибулярное ядро Дейтерса
в первой группе ни у одной крысы. У животных подгруппы Б, с быстро
регрессировавшими послеоперационными неврологическими нарушениями, в двух
случаях очаг повреждения захватывал область моторного ядра лицевого нерва и
гигантоклеточного ретикулярного ядра; еще в одном наблюдении этот очаг не был
обнаружен вовсе. В подгруппе А, у крыс, с отсутствовавшими после операции
видимыми изменениям позы и двигательной активности, в 5 случаях имело место
попадание коагулирующего электрода в область вентрального и дорсального кохлеарного
ядер (рис. 3.5,Б,2), в одном - в левую ножку мозжечка. У одной крысы очаг
разрушения не определялся.
Во второй, «осложненной», группе у 8 из 9 крыс
обнаружено точное попадание коагулирующего электрода в левое вестибулярное ядро
Дейтерса. У одной крысы морфоконтроль не проводился. При этом объем и характер
разрушения варьировал у разных животных. Отмечены случаи локального (рис.
3.5,Б,1) и довольно обширного разрушения вестибулярного ядра с большой
протяженностью в рострокаудальном направлении (рис. 3.5,Б,3). У двух животных
наблюдалось полное замещение области разрушения глиальным рубцом. Важным
представляется тот факт, что течение послеоперационного периода по
неврологическим показателям не зависело только от объема стволового разрушения.
Так, у животного с самым обширным повреждением ядра Дейтерса (рис. 3.5,Б,3)
обнаружена достаточно быстрая (через полторы недели) и полная компенсация
нарушенных функций. Такими же были сроки редукции неврологических отклонений и
в ряде наблюдений с ограниченными разрушениями. В то же время у некоторых крыс
с аналогичным или даже меньшим объемом повреждения (рис. 3.5,Б,1) отмечались
грубые, не компенсируемые в течение полутора месяцев, поведенческие нарушения.
Судя по всему, последствия локального стволового повреждения определяются
многими факторами, к числу которых относятся реакция ткани и сосудов мозга на
электротравму, целостные церебральные реакции на повреждение и др.
Из семи крыс, погибших во время или в разные
сроки после операции, патоморфологические исследования удалось провести лишь в
двух случаях. Если в одном из них имело место довольно грубое повреждение ядра
Дейтерса, то во втором точное локальное попадание коагулирующего электрода в
левое ядро Дейтерса сопровождалось сравнительно небольшим объемом разрушения
(рис.3.5,Б,2).
Сходство особенностей гистологической картины
повреждения у этого животного и животных из второй группы является еще одним
подтверждением нашего вывода о том, что тяжесть послеоперационного состояния не
сводится только к локализации и объему разрушения, но определяется,
по-видимому, вторичными дисгемическими нарушениями и особенностями протекания
системных адаптивно-компенсаторных процессов, которые обусловлены, в частности,
индивидуальными особенностями ЦНС.
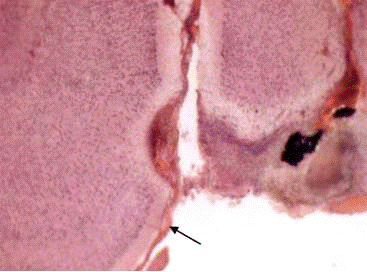
Рис.3.5. Фронтальные срезы мозга крыс при
стволовом повреждении: 1 -вариант с локальным разрушением ВЯД; 2 - повреждение
ствола с деструкцией кохлеарных ядер; 3 - обширное разрушение ствола на этом же
уровне (показано стрелками)
Сопоставление результатов наших
экспериментальных исследований с данными литературы свидетельствует об
определенном сходстве последствий повреждения ствола мозга у крыс с
проявлениями стволовой патологии у человека, что обусловлено определенной
универсальностью организации звена стволовой регуляции этих функций у
млекопитающих. Известно, что латеральное вестибулярное ядро Дейтерса, наряду с
красным ядром, является частью системы, регулирующей позу и двигательные
рефлексы (Шмидт, Тевс, 1985; Лиманский, 1987; Baskai
et al.,
2002; Krutki
et al.,
2003). От ядра Дейтерса берет начало вестибулоспинальный тракт, оказывающий
возбуждающее влияние на альфа- и гамма-мотонейроны разгибателей и тормозное -
на мотонейроны сгибателей. Одностороннее разрушение ядра Дейтерса приводит к
повышению тонуса мышц-сгибателей на ипсилатеральной стороне, в основном, на
уровне шейного и грудного отделов. Поэтому специфическое изменение позы (наклон
головы к туловищу), выявляемое у животных с точечным попаданием коагулирующего
электрода в латеральное вестибулярное ядро Дейтерса, характерно и для клиники
поражения стволовых вестибулярных ядер человека (Сепп c
соавт., 1950). Причем, легкие и кратковременные нарушения позы в виде наклона
головы к туловищу под углом 30 градусов, отмечаемые у животных первой группы
при попадании коагулирующего электрода в область кохлеарных ядер, ножку
мозжечка и др. структуры ствола, могут быть объяснены тоническими влияниями от
разрушенных ядер моста, имеющих афферентные связи с ядром Дейтерса (Лиманский,
1987), затеканием тока в структуры ядра Дейтерса (Егоров, Кузнецова, 1972), а
также вторичными реакциями на механическую и электрическую травму ствола
(локальная зона отека мозговой ткани, локальное нарушение микроциркуляции)
(Смирнов, 1949), которые не обнаруживаются при используемом методе окрашивания
срезов мозга.
Причиной появления у животных винтообразных
движений является, по-видимому, снятие фазного тонического контроля
антигравитационной мускулатуры, который осуществляется через
вестибулоспинальный тракт и нарушается при повреждении ствола на уровне ядра
Дейтерса (Магнус, 1962; Котляр, Шульговский, 1979; Basso
et al.,
2002; Uno
et al.,
2003). Невозможность удержания на гладкой поверхности может быть объяснена
возникновением системного головокружения, характерного для застойных очагов
возбуждения в вестибулярном анализаторе (Благовещенская, 1990). Появление
сукровичных подтеков, вероятно, можно объяснить наличием вестибулярных проекций
к ядрам глазодвигательных мышц (McMasters
et al.,
1966)/
В клинике опухолей задней черепной ямки, равно
как и при других формах поражения ствола мозга человека с вовлечением в
патологический процесс вестибулярных образований, указанные неврологические
нарушения также проявляются в качестве ведущих, хотя и в менее "чистом
виде" по сравнению с животными. К числу характерных и аналогичных
экспериментальным симптомов можно отнести частые головокружения с вынужденной
позой головы и туловища, шейные тонические реакции с поворотом головы,
гиперкинезы стволового происхождения. Разновидностью последних является
спастическая кривошея. Отмечались также нарушения статики и походки
(вестибулярная атаксия) (Сепп с соавт., 1950; Чернышев, 1983; Благовещенская,
1990).
Наблюдение за состоянием животных после
электролитической стволовой коагуляции и распределение их по трем группам в
зависимости от особенностей динамики их состояния и исходов также указывает на
определенные аналогии с вариантами послеоперационного течения у
нейрохирургических больных после удаления стволовой опухоли (Шарова, 1991,
1993).
Эти обстоятельства свидетельствуют о
правомочности использования предложенной модели локального электролитического
повреждения ствола мозга для изучения адаптивно-компенсаторных перестроек ЦНС
при острых стволовых повреждениях - в том числе и для выяснения некоторых
неясных аспектов формирования таких реакций у человека.
.3
Анализ зависимости послеоперационного течения от дооперационных
особенностей
поведения животных
Уже предварительные морфо-функциональные
сопоставления обследованных нами животных позволяют предположить значимость
специфики протекания церебральных адаптивно-компенсаторных реакций и
индивидуальных особенностей ЦНС в исходе рассматриваемой экспериментальной
патологии. В исследованиях многих авторов показано, что устойчивость человека и
животных к различного рода неблагоприятным воздействиям зависит от типа высшей
нервной деятельности (Айрапетянц с соавт, 1980), эмоциональности и подвижности
(Зухарь, 1985; Саркисова с соавт., 1992), а также доминантности полушарий
(Брагина, Доброхотова, 1988). Широко известна активная роль лимбической системы
в приспособительной деятельности мозга (Вейн, Соловьева, 1973; Ониани, 1980).
Причем, электрографические признаки активизации отдельных ее составляющих на
определенных этапах адаптивного процесса при остром стволовом повреждении
выявлены у больных после удаления опухоли ствола (Шарова с соавт., 1991; 1993;
1995).
Все эти обстоятельства побудили нас к детальному
сопоставлению дооперационных эмоционально-поведенческих особенностей животных с
динамикой их функционального состояния и исходом в условиях локального
стволового повреждения.
До коагуляции ствола мозга 26 из 28 крыс были
протестированы в открытом поле. У животных каждой из выделенных
послеоперационных групп была проведена целостная оценка исходной двигательной
активности, а также отдельных ее составляющих: горизонтальной и вертикальной
активности - с их последующим ранжированием (см. главу 2).
Согласно результатам статистических
сопоставлений горизонтальной двигательной активности животных до операции и послеоперационного
исхода, имеется тенденция (p
= 0.07) к ухудшению исхода по мере возрастания фонового уровня этой активности
(табл.3.2).
В группе с неосложненным послеоперационным
течением приблизительно равномерно представлены все степени горизонтальной активности;
среди особей с осложненным течением преобладали животные со средней и высокой
активностью; в группе с летальным исходом самый большой, по сравнению с другими
группами, процент животных с очень высокой горизонтальной активностью.
Таблица 3.2. Распределение животных с разным
уровнем горизонтальной двигательной активности до операции по исходам после
стволового повреждения
Исход
горизонтальная
Неосложненный
(11)
Осложненный
(8)
Летальный
(7)
Низкая
1
(10%)
-
1
(14%)
Средняя
3
(27%)
3
(28%)
- 4
(36%)
4
(50%)
2
(29%)
Очень
высокая
3
(27%)
1
(13%)
4
(57%)
Примечание: В скобках - процент от количества
животных в группе.
Любопытные тенденции выявлены для показателей
вертикальной активности. Наименьшее число вертикальных стоек в фоне присуще
животным с неосложненным послеоперационным течением (50%). Оно значимо меньше,
в частности, чем у крыс с осложненным послеоперационным течением, для которых
характерен умеренно высокий уровень вертикальной активности (табл. 3.3). Но при
этом низкая вертикальная активность в фоне преобладала и в группе с летальным
исходом.
Рис. 3.6. График взаимоотношений вертикальной и
горизонтальной активности
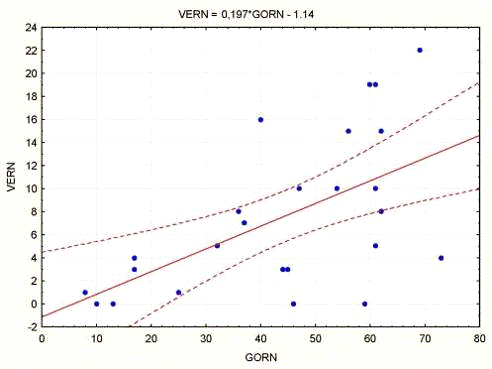
Интересно, что горизонтальная и вертикальная
составляющие двигательной активности оказались связанными между собой линейно с
К кор. = 0.58 (при Р < 0.002) по уравнению регрессии
активность
верт. = 0.197 акт. гор. - 1.14.
Таблица 3.3. Ранжированное распределение
животных по исходам в зависимости от уровня вертикальной активности.
Исход Вертикальная
неосложненный
осложненный
летальный
Сумма
по рядам
Нулевая
2
(50%)
1
(25%)
1
(25%)
4
Низкая
7
(44%)
4
(25%)
5
(31%)
16
Высокая
2
(33%)
3
(50%)
1
(17%)
6
Сумма
по колонке
11
(42%)
8
(31%)
7
(27%)
26
Примечание: в скобках - процент от общего
количества животных с данным видом
активности
Нами были исследованы также соотношения между
общей двигательной активностью животных до операции и исходом. Общая
двигательная активность определялась как суммарное значение вертикальной,
горизонтальной активности и количества вращений.
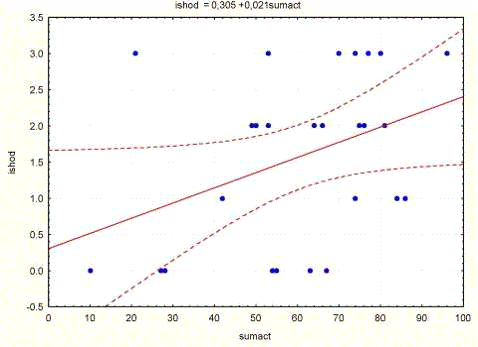
Рис.3.7. График, отражающий положительную
корреляционную связь между суммарной активностью и исходом
Выявлено наличие положительной корреляции между
этими показателями: коэффициент линейной корреляции равен 0.39 при р = 0.05,
тогда как ранговый коэффициент корреляции Спирмана К = 0.34 при р = 0.09. Из
графика, приведенного на рис. 3.7 следует, что ранг исхода животных (0,
1неосложненный, 2 - осложненный с выживанием, 3 - летальный) увеличивается по
мере роста их дооперационной суммарной двигательной активности. Эти соотношения
можно, по-видимому, рассматривать как отражение связи между исходным уровнем
беспокойства животного (а не уровнем исследовательской активности) и возможностью
компенсации: чем выше уровень беспокойства, тем менее благоприятен исход
патологического процесса.
Не было выявлено убедительной связи между
"рукостью" животных, обусловленной,
скорее всего, асимметрией коркового конца двигательного
анализатора,
и послеоперационным исходом. Вместе с тем, отмечено
определенное
соответствие между предпочитаемым направлением движения
крыс
в открытом поле и исходом: 7 из 12 животных с неосложненным
течением
(т. е.64%) предпочитали до операции движение в открытом поле
вправо,
тогда как 6 из 7 погибших (89%) - влево (табл. 3.4).
Таблица 3.4. Распределение животных с
изолированным стволовым повреждением по послеоперационным исходам в зависимости
от предпочитаемого направления движения и эмоционального фона до операции
N Группы
Исход
Число
оперированных животных
Число
тестированных животных
Число
животных с повышенной эмоциональностью
Предпочитаемое
направление движения правое левое
1
Неосложненный
12
11
7
(64%)
7 (64%)
4 (36%)
II
Осложненный
9
8
6
(75%)
3 (38%)
5 (62%)
III
Летальный
7
7
6
(89%)
1 (11%)
6 (89%)
Всего:
28
26
19
11
15
В литературе этот вид моторной асимметрии
связывают преимущественно с асимметрией в пределах нигростриарной системы,
доказательством
чего, по мнению авторов, служит изменение ротации животных
под действием препаратов, влияющих на метаболизм дофамина
(Бианки
с соавт., 1982; Вартанян, Клементьев, 1991; Glick
et al.,
1977; Strauss
et al.,
1983). Таким образом, считается, что характер пространственно-моторного
предпочтения
определяется, в основном, дисбалансом между двумя нигростриарными
системами и зависит от их взаимодействия с корковыми
отделами
обоих полушарий. Существует мнение, что в проявлении разных
видов
моторной активности ведущим является сенсорный компонент реакции
(Удалова,
Михеев, 1982). Не отмечено корреляции пространственно-моторного
предпочтения
с «рукостью» животных (Мельников с соавт., 2001), что может
быть
обусловлено разными механизмами формирования этих видов
асимметрий
(Бианки, 1980; Наливаева с соавт., 1993).
Даже самое общее сопоставление эмоционального
фона животных до операции
повреждения ствола с исходом указывало на наличие связи между
этими
характеристиками (табл.3.4): число крыс с исходно повышенной
эмоциональностью
нарастало и достигло максимума в группе с летальным
исходом.
Более детальный статистический анализ подтвердил наличие этой,
хотя
и нелинейной (немонотонной) связи (р = 0.08).
Таблица 3.5 Распределение крыс по
исходам в зависимости от исходного уровня эмоциональности
-Эмоциональность
Исход
0
1
2
3
0
4 (66.7%)
1
(9.1%)
2
(33.3%)
0
1
0
1
(9.1%)
1
(16.7%)
2
(66.7%)
2
1 (16.7 %)
4
(36.4%)
2
(33.3%)
1
3
1
5
1
Примечание: ранги 0 и 1 в колонке
исходов соответствует подгруппам А и Б с неосложненным исходом
Как следует из таблицы 3.5, повышение вероятности
летального исхода связано с повышением уровня эмоциональности с 0 до 1 (то есть
с фактом наличия или отсутствия эмоциональных реакций). При увеличении уровня
эмоциональности (ранги 2 и 3) вероятность неблагоприятных исходов (уровня
больше 1) статистически неразличимы.
Изучение соотношений между наличием
эмоциональных реакций у животных до операции и компонентами их двигательной
активности на основе ранговых корреляций Спирмана показало, что вертикальная
активность и число вращений связаны с эмоциональными реакциями в более высокой
степени (R = 0.32), чем
горизонтальная (R = 0.13).
Следует отметить, что эти коэффициента статистически недостоверны (р > 0.10)
- отчасти, вероятно, из-за недостаточного числа наблюдений.
Послеоперационное наблюдение за состоянием
животных в динамике выявило наличие специфических эмоциональных реакций,
различающихся по своим характеристикам в группах с разным послеоперационным
исходом. Так, при неосложненном послеоперационном течении только у одной крысы
из подгруппы А, при отсутствии нарушений позы и движений, наблюдалась
отчетливая эмоциональная реакция - вокализация - на тактильное раздражение. В
группе с осложненным послеоперационным течением у всех 9 животных отмечались
выраженные эмоциональные реакции: сильный писк (спонтанный или при тактильных
раздражениях), яркая оборонительная реакция.
Эти реакции у четырех особей были выражены
волнообразно (на 2-3-и сутки, 13-ые сутки и 22-23-и сутки после операции), а у
трех животных эмоциональные реакции наблюдались длительное время: со вторых по
десятые сутки и с первых по двадцать четвертые сутки (рис.3.8, Б,В).
Описанные эмоциональные проявления были присущи
как животным с наличием эмоциональных реакций до операции, так и животным, у
которых такие реакции не наблюдались. У четырех крыс отмечена тенденция к
совпадению по времени появления эмоциональных реакций и положительных изменений
неврологического статуса.
В Рис. 3.8. Появление эмоциональных реакций у крыс
с летальным исходом (А) и у крыс с осложненным течением послеоперационного
периода (Б, В). По оси ординат - сутки после операции; по оси абсцисс
-формализованные показатели эмоциональных проявлений (1 - слабая оборонительная
реакция; 2 - сильная оборонительная реакция; 3 - агрессивно-оборонительная
реакция.
Из семи животных третьей («летальной») группы
четыре особи погибли на операционном столе. У двух из трех оставшихся крыс
накануне летального исхода отмечены изменение в эмоциональной сфере, несколько
отличавшееся по характеру от реакций крыс второй группы: агрессивность,
вокализация, а также повышение мышечного тонуса при взятии в руки (рис.3.8,А).
Сравнивая животных в группе с изолированным
повреждением ствола до и после операции, следует отметить увеличение количества
крыс с
наличием
эмоциональных реакций. Но это увеличение было статистически недостоверным (по
критерию %2 р=0.5), (рис.3.9).
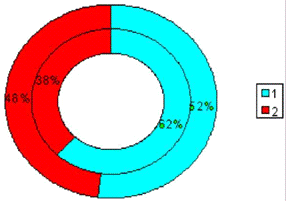
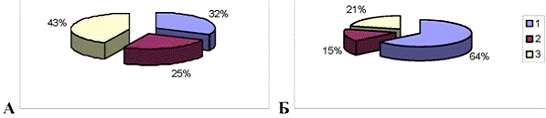
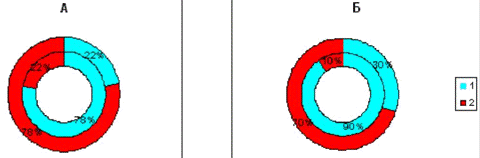
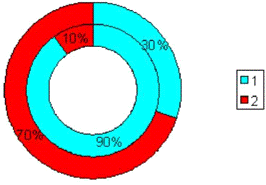
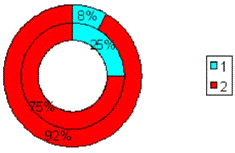
Рис.3.9. Диаграмма, показывающая изменение
количества эмоциональных животных в группе с изолированным стволовым
повреждением (наружный круг) по сравнению с количеством эмоциональных крыс до
операции (внутренний круг). 1 - спокойные животные; 2 - эмоциональные животные.
Проведенный анализ позволяет отметить, что при
разных типах послеоперационного течения после локального повреждения ствола
мозга имеет место мобилизация ресурсов ЦНС путем активации структур лимбической
системы, выражающееся в появлении эмоциональных реакций в критические моменты
компенсаторного процесса. Наблюдается также разновариантность эмоциональных
реакций в случае разных исходов, что, возможно, отражает особенности
функционирования лимбических структур в постоперационный период. Эти
обстоятельства обосновывают целесообразность проведения более детального
исследования роли отдельных лимбических образований в послеоперационных
адаптивно-компенсаторных реакциях - в частности, с привлечением
электрофизиологических методов.
Глава 4. Электрофизиологические исследования у
животных
с локальным стволовым повреждением
Электрическая активность головного мозга (ЭА),
дополненная современными методами математического анализа является
информативным и объективным показателем функционального состояния мозга
человека и животных (Павлыгина, Соколов 1983; Русинов c
соавт., 1987; Русинова, 1989). Если изменения вызванных потенциалов в ответ на
повреждение ствола мозга животных в определенной степени изучены (Незлина
1957), то данные о компенсаторных перестройках спонтанной ЭА коры больших
полушарий и других заинтересованных структур требуют существенного дополнения.
Цель настоящего раздела работы - динамический
анализ пространственно-временной организации электрической активности мозга
крыс при остром локальном повреждении ствола для изучения развивающихся при
этом церебральных процессов компенсации (либо декомпенсации), с уточнением
характера включения в них ОФК и гиппокампа.
У 8 животных (7 особей - самцы белых беспородных
крыс, 1 - самец линии Wistar)
в 27 опытах регистрировали спонтанную электрическую активность головного мозга
до и после левостороннего электролитического разрушения латерального ВЯД.
Вживление электрода, через который производилось
электролитическое разрушение ствола на уровне ядра Дейтерса, производилось по
схеме, описанной в главе 2.
Регистрирующие платиновые электроды вживляли в
симметричные орбитофронтальные и соматосенсорные области коры, в поле СА1
правого и левого гиппокампа и интактное ВЯД справа - с последующим
морфологическим контролем локализации электродов (глава 2).
Биоэлектрическая активность мозга крыс до
разрушения ствола мозга.
Так как в литературе мы не встретили детального
описания пространственно-временной организации биоэлектрической активности
мозга крыс, основанном на методах обсчета, применяемых в клинике, остановимся
на этом вопросе подробнее.
Визуальная оценка рисунка фоновой ЭА выявила
чередование эпизодов синхронизации и десинхронизации, охватывающих все
регистрируемые области. Причем, у 5 из 8 животных характерным было
превалирование десинхронной части над синхронной в 1.3 - 3.3 раза (рис.4.1 А1).
Лишь в двух наблюдениях отношение длительности десинхронной ЭА к
синхронизированной составило 0.7 и 0.9.
Отмечались региональные различия амплитуды
сигнала: более низкая - в орбитофронтальных областях коры и интактном стволе
при максимальных ее величинах и экзальтации составляющих ЭА в гиппокампе (рис.
4.1 А).
В целом биоэлектрическая активность
характеризовалась полиритмией: во всех отделах превалировали тета-составляющие
в сочетании с группами альфа- и бета-колебаний. Тета-активность имела наиболее
ритмичный характер в гиппокампе. Элементы синхронизации ЭА отмечались и на
десинхронных участках записи.
Спектральный анализ ЭА позволил подтвердить и
дополнить эти наблюдения. На спектрах мощности десинхронной составляющей ЭА
выявлялись, как правило, два пика: в дельта-диапазоне и в тета-полосе, на
частоте 6-7 Гц (рис. 4.1 А). Причем, частотная разница доминирующих пиков между
исследуемыми областями отсутствовала, либо была выражена без устойчивой
закономерности. Качественные различия спектрального состава между синхронной и
десинхронизированной частями записи были выражены нерезко, хотя мощность всех
частотных составляющих была больше на синхронных реализациях - особенно в
тета-диапазоне. В некоторых случаях несколько менялись (подчеркивались)
различия в мощностных соотношениях пиков.
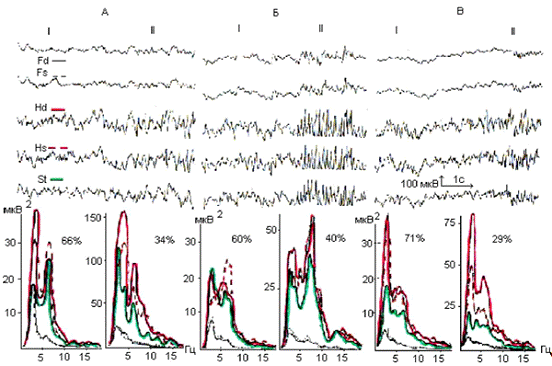
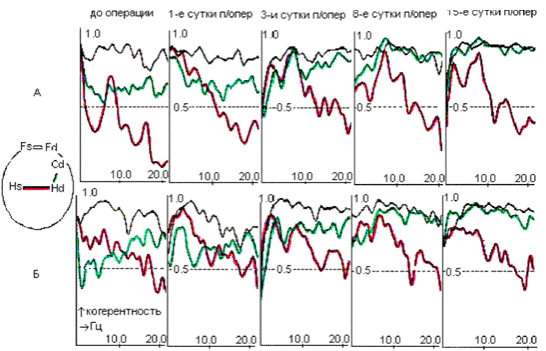
Рис. 4.1. Электрическая активность и спектры
мощности симметричных областей ОФК (Fd;
Fs), поля СА1
гиппокампа (Hd; Hs)
и ствола (St) мозга c
неосложненным послеоперационным течением до (А), в первые (Б) и тридцатые (В) сутки
после электролитической коагуляции латерального вестибулярного ядра Дейтерса
слева: I - десинхронная
составляющая; II - синхронная составляющая
При когерентном анализе десинхронных реализаций
ЭА были установлены определенные соотношения сочетанности биопотенциалов между
разными регионами мозга (рис. 4.2, I; 4 AI).
Так, у пяти животных максимальные значения когерентности ЭА (порядка 0.6 - 0.8)
были характерны для симметричных отделов орбитофронтальной коры (рис. 4.2 AI;
4.5
AI)
Величины когерентности, равные 0,4-0,6
отмечались между биопотенциалами симметричных соматосенсорных и гиппокампальных
областей мозга, а также между орбитофронтальной и соматосенсорной корой в
пределах одного полушария (рис. 4.2 I
Б,В; 4.5 AI). Связям
ОФК с гиппокампом были присущи более низкие значения (0.3-0.4).
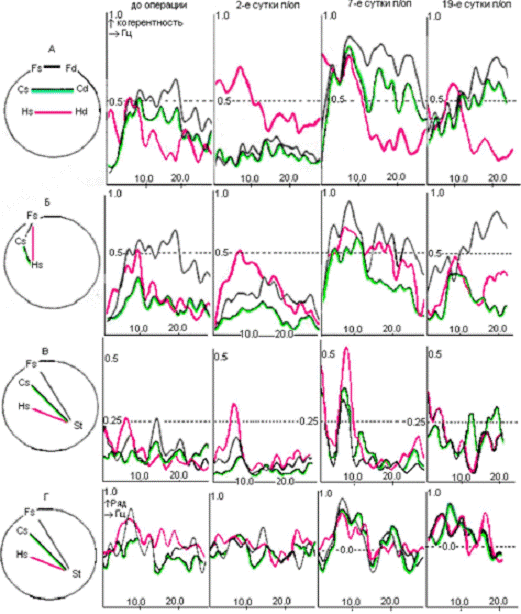
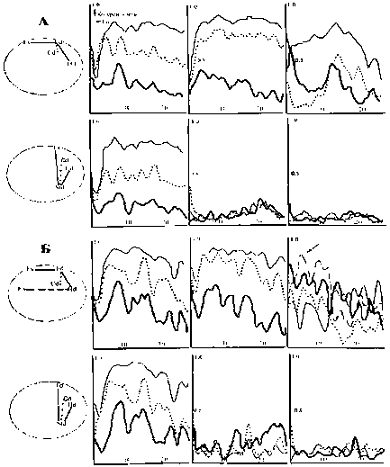
Рис.4.2. Динамика когерентных (А, Б,
В) и фазовых (Г) отношений десинхронной составляющей ЭКоГ у крысы с
повреждением ствола при неосложненном послеоперационном течении. Обозначения областей
регистрации как на рис 4.1
Наконец, минимальная когерентность (порядка
0.1-0.2) определялась между ЭА интактного ствола и всеми остальными
регистрируемыми зонами мозга (рис. 4.2 I,B;
4.5 AI).
Интересно, что именно у этих пяти животных общая длительность десинхронной
части ЭА превосходила синхронную. Для особей с обратным соотношением
десинхронной и синхронной составляющей (т.е. при более низком, по-видимому,
уровне бодрствования) специфичным было снижение сочетанности биопотенциалов
симметричных отделов ОФК по сравнению с симметричными соматосенсорными или
гиппокампальными зонами, а также орбитофронтальной и сенсомоторной корой в
пределах одного полушария.
К числу особенностей синхронизированной
составляющей ЭА (рис. 4.3 I; 4.5 БГ) относится, прежде всего, общая тенденция к
более высоким (по сравнению с десинхронной частью записи) значениям
когерентности в широком частотном диапазоне.
Рис. 4.3. Динамика спектров
когерентности синхронизированной составляющей электрической активности у
животного с повреждением ствола при неосложненном послеоперационном течении.
Обозначения областей регистрации как на рис 4.1
Это было особенно отчетливо выражено на внутри-
и межполушарных гиппокампальных связях. В некоторых случаях сочетанность
симметричных гиппокампальных зон была больше лобных, тогда как на десинхронной
части записи - наоборот. У некоторых животных зональные соотношения
когерентности изменялись за счет усиления стволово-полушарного взаимодействия.
Рисунок спектров когерентности симметричных
отделов ОФК, а также ствола с другими образованиями имел по большей части
"шумовой" характер. В ряде случаев отмечалось наличие нескольких
пиков: на частотах 6, 7, реже -8 Гц (рис.4.2, 4.3, 4.5 I). У трех животных было
выявлено повышение когерентных связей на частотах бета-диапазона (около 12 или
14-18 Гц).
При попытке исследования зависимости
особенностей ЭА крыс от моторной доминантности полушарий (Е.И.Микляева, 1989)
была отмечена тенденция к существованию связи между "рукостью"
животного и мощностью биопотенциалов (р = 0.08; коэффициент сопряженности 0.40
по точному критерию Фишера): мощность ЭА в корковых (прежде всего
сенсомоторной) областях, как правило, превалировала в "доминантном"
(контралатеральном предпочитаемой передней лапе) полушарии. Значения же
внутриполушарных когерентностей (особенно между орбитофронтальной и
сенсомоторной корой) достоверно преобладали в "субдоминантном"
полушарии - более отчетливо на десинхронных участках записи ( р = 0.035;
коэффициент сопряженности 0,38 по точному критерию Фишера). Это обстоятельство
было довольно неожиданным, учитывая данные литературы о больших значениях этого
показателя в доминантном полушарии у человека (Болдырева, 1978; Жаворонкова,
1990).
Фазовые спектры ЭА на десинхронных участках
записи имели шумовой характер с попеременной сменой опережающей области - в
зависимости от частотного диапазона. При этом в парах со стволом
"лидирующей" (особенно на частотах ниже 20 Гц) чаще оказывалась
именно эта структура (рис. 4.2Г); в парах симметричных отведений
"опережало" по большей части правое полушарие, а во внутриполушарных
связях - менее глубинная зона или область, лежащая фронтальнее. Специфичным же
для синхронизированных участков ЭА является тот факт, что на внутриполушарных
фазовых спектрах чаще " лидировали" более глубинно расположенные
области.
После стволовой электролитической коагуляции
животные разделились по трем вариантам послеоперационнй динамики состояния,
описанным нами ранее (глава 3). У двух особей с легким, быстро (до трех суток)
регрессировавшим наклоном головы, послеоперационное течение было расценено как
неосложненное. По данным морфоконтроля (через 20 и 35 дней после коагуляции) у
этих крыс имело место частичное разрушение в кохлеарном или ретикулярном ядрах
ствола. В послеоперационной динамике ЭА у этих особей были выявлены следующие
особенности по сравнению с фоном.
. Изменились соотношения длительностей
десинхронной составляющей к синхронной: уменьшилась доля десинхронной части
записи - либо сразу после операции (1-е сутки) (рис. 4.1 Б), либо к 7-м суткам,
после резкого первоначального ее увеличения. Приближение этого показателя к
фоновому уровню было отмечено на 19-е и 30-е сутки после стволового повреждения
(рис.
4.1 В).
2. В динамике ЭА и ее спектров мощности выявлено
нерезкое уменьшение общей мощности сигнала и выраженности составляющих
дельта-диапазона при относительном усилении тета-активности в диапазоне 6-7 Гц
в 1-2 сутки после повреждения (в обоих случаях, как на синхронной, так и
десинхронной частях записи) (рис. 4.1 Б с последующей нормализацией
спектральных характеристик ЭА (рис. 4.1В Наиболее отчетливо изменялась
активность гиппокампа, особенно на синхронном участке записи: наблюдалась
экзальтация колебаний и появление эпилептиформных черт, сохраняющихся в
процессе всего наблюдения (рис.4.1Б В
3. Наиболее выраженными и динамичными были
изменения пространственного взаимодействия биопотенциалов. На десинхронной
составляющей ЭА (рис. 4.2 II)
в 1-2-е сутки после разрушения ствола наблюдалось снижение большинства
когерентностей ЭА относительно фонового уровня в широком частотном диапазоне -
за исключением гиппокампальных связей. Наряду с этим отмечено избирательное
повышение некоторых из них. Прежде всего, это относится к уровню когерентности
на частоте около 6 Гц между стволом и гиппокампом (с фазовым опережением
ствола), между симметричными гиппокампальными областями и, в меньшей степени,
между гиппокампом и корковыми зонами (орбитофронтальная и сенсомоторная кора) в
пределах полушарий - без четкой латерализации. Причем, пики повышенной
сочетанности на спектрах этих когерентностей имели более очерченный по
сравнению с фоном характер. К 7-м суткам после стволовой деструкции связь на
частотах 6-7 Гц приобретала более генерализованный характер (рис. 4.2 III).
Она усиливалась между интактным стволом и гиппокампом, стволом и корковыми
зонами; охватывала также большинство исследуемых корковых областей - как между
симметричными, так и унилатеральными орбитофронтальными и сенсомоторными
зонами. Взаимодействие же между симметричными областями гиппокампа несколько
ослабевало. В дальнейшем отмечалась тенденция к "нормализации"
показателей когерентности ЭА (19-е сутки) (рис. 4.2 IY),
либо снижению их по сравнению с фоновым уровнем (30-е сутки). Значимость
выявленных изменений когерентности была подтверждена с помощью статистической
обработки по программе В.Г.Воронова (2000).
На синхронных участках ЭА в 1-2-е сутки после
стволовой коагуляции стволово-гиппокампальное взаимодействие также несколько
усиливалось, но в области более медленных (около 2 Гц) или высоких (8, 17 Гц)
ритмов (рис. 4.3 II).
При этом избирательное повышение межгиппокампальной (в обоих случаях),
корково-гиппокампальной, а также внутриполушарной лобно-сенсомоторной
синхронизации выявлялось на частоте 4-5 Гц. На этом фоне происходило увеличение
сочетанности биопотенциалов между симметричными орбитофронтальными областями: в
широком частотном диапазоне в одном случае (рис. 4.3 III), на частотах 10-12 Гц
- в другом. К 7- 19 суткам после деструкции структура всех рассматриваемых
когерентных отношений практически нормализовалась (рис. 4.3 IY)
Таким образом, у животных с неосложненным
послеоперационным течением разрушения на уровне ствола в условиях
морфологической сохранности ВЯД вызывает, тем не менее, снижение его тонизирующих
влияний на корковые отделы. При этом в более коротких (по сравнению с фоном)
эпизодах бодрствования модифицируется функциональная связь ствола с
гиппокампом, что приводит к генерализованному усилению межгиппокампальной,
корково-гиппокампальной и корково-корковой моночастотной (6-7 Гц)
синхронизации. Это указывает на формирование единой системы повышенной
функциональной активности при ведущей роли ствола. Во время удлинившихся
эпизодов засыпания (синхронные участки ЭА) стволово-гиппокампальное взаимодействие
также усиливается, но по патологически низким либо, напротив, высоким ритмам.
Может наблюдаться, в частности, кратковременное (1-3 сутки после операции)
нарастание синхронизации на частоте 4-5 Гц между симметричными областями
гиппокампа, гиппокампом и другими исследуемыми структурами мозга,
соответствующее эпизодам угнетения стволовой активности и имеющее в данной
группе животных обратимый характер.
В трех наблюдениях послеоперационное течение
животных расценивалось как осложненное. Длительное время (до 10 суток)
сохранялся наклон головы в оперированную сторону. В ранние сроки после
стволового повреждения все крысы не могли удерживаться на гладкой поверхности и
заваливались на бок.
В одном наблюдении имело место появление
винтообразных движений. Морфоконтроль выявил во всех случаях попадание
коагулирующего электрода в левое ВЯД.
Для динамики их ЭА было характерно следующее:
1. Как и в случаях с неосложненным
послеоперационным течением, изменялись временные соотношения десинхронной и
синхронной составляющей. В двух случаях, где до стволовой коагуляции
десинхронная часть значительно (более чем в три раза) преобладала над
синхронной, отмечено явное увеличение длительности последней (вплоть до
инверсии этих соотношений), сохранявшееся на протяжении трехнедельного
исследования. У крысы с исходно превалирующей по времени синхронной ЭА
произошло, напротив, относительное увеличение длительности десинхронной
составляющей.
2. Динамика рисунка ЭА и спектров мощности
в общих чертах повторяла послеоперационные изменения, имеющие место при
неосложненном течении. Имела место экзальтация и даже эпилептизация
гиппокампальной активности (особенно на десинхронных участках записи); большая
пароксизмальность синхронных фрагментов; динамичность спектрального состава (период
относительного нарастания мощности медленных волн сменялся большей
очерченностью и величиной тета-пика - с последующей тенденцией к нормализации
спектрограммы). Однако если при самом легком из осложненных послеоперационных
вариантов весь этот цикл изменений заканчивался к 16-м суткам, то в двух
других, более тяжелых случаях нейродинамические этапы были сдвинуты и затянуты
по времени (по сравнению с неосложненным течением), достигая своего максимума к
9-10 суткам.
3. Динамика межцентральных отношений (рис.4.4)
десинхронной составляющей ЭА также была сходна с изменениями при неосложненном
послеоперационном течении: тенденция к усилению гиппокампальных связей на
частоте 6-7 Гц с постепенной генерализацией этих влияний на другие зоны мозга,
максимально выраженной на 4-7-е сутки после операции. Причем, чем тяжелее
состояние животного, тем длительнее была выражена такая генерализация.
Характерным для данной группы крыс было то, что повышение когерентности на
частоте 6-7-Гц имело место и на синхронном участке ЭА.
Рис.4.4 Динамика спектров когерентности
десинхронной (А) и синхронной (Б) составляющих ЭА у крысы с повреждением ствола
при осложненном послеоперационном течении. Обозначения областей регистрации как
на рис 4.1
При этом "лидерство" ствола в запуске
описываемой реакции у крыс этой группы выражено менее отчетливо по сравнению с
"неосложненной" группой. Так, у одного животного начальным (1-е сутки
после операции) было повышение межполушарной орбитофронтальной когерентности
десинхронной составляющей ЭА на частоте 6,5 Гц. У другого в 1-е сутки после
стволовой коагуляции имело место усиление межполушарной орбитофронтальной связи
(с нерезким пиком 7 Гц) - наряду с повышением стволово-полушарной когерентности
в тета-диапазоне (при фазовом опережении ствола) на синхронных фрагментах ЭА.
Стволово-полушарное взаимодействие десинхронной составляющей в тета-диапазоне
было при этом ослаблено. У третьей крысы формировались как бы два встречных,
"запускающих" синхронизацию по близким частотам, потока:
полушарно-гиппокампальный (на частоте 7 Гц), с одной стороны, и
стволово-гиппокампальный (на частоте 6 Гц) - с другой.
Таким образом, в случаях осложненного течения,
при существенном начальном ослаблении активирующих стволовых влияний, длительному
усилению внутри- и межгиппокампальной синхронизации биопотенциалов на частоте
6-7 Гц с последующей генерализацией этого феномена может предшествовать
повышение не только стволово-гиппокампальных связей, но и когерентности между
симметричными орбитофронтальными либо полушарно-гиппокампальными областями в
широком или избирательном (6-7 Гц) частотном диапазоне.
Следует отметить, что основные черты
послеоперационной нейродинамики были присущи и крысе Wistar,
отнесенной к группе с осложненным течением послеоперационного периода.
У двух животных разрушение ВЯД привело к
летальному исходу, наступившему в одном случае на пятые, в другом - на 17-е
сутки после стволовой коагуляции. Наиболее демонстративной была динамика
состояния особи с быстрым летальным исходом. Сразу после операции выявились
грубые стволовые неврологические симптомы: нарушение движений, заваливание на
левый бок, вытягивание конечностей. К пятым суткам проявилась цианотичность
конечностей на фоне резкого снижения веса и появилось нарушение дыхания.
Послеоперационная динамика биоэлектрической
активности этой крысы характеризовалась следующими особенностями (рис.4.5):
1. К третьим суткам резко (примерно в три
раза) увеличилась относительная длительность десинхронной составляющей ЭА по
сравнению с фоном, сохраняющаяся (хоть и в меньшей степени) вплоть до
летального исхода.
2. Изменения рисунка ЭА на третьи сутки после
разрушения ствола существенно не отличались от таковых у выживших животных. Они
заключались в появлении эпилептиформных черт в активности, главным образом,
гиппокампа и сенсомоторной коры. Кроме того, имела место слабая очерченность
синхронизированных участков ЭА и их локализованность: вместо генерализованных
вспышек - фрагменты ритмической активности в сенсомоторной коре и гиппокампе.
Лишь на пятые сутки выявились отчетливые, специфические сдвиги ЭА: резкое
снижение уровня биопотенциалов и уплощение рисунка на десинхронной составляющей
записи, чередующееся с высокоамплитудными пароксизмами эпилептиформной
активности (комплексы "острая-медленная волна", веретена частых
колебаний).
Рис.4.5. Динамика спектров
когерентности десинхронной (А) и синхронной (Б) составляющей ЭКоГ животного с
летальным исходом после стволовой коагуляции. Обозначения областей регистрации
как на рис 4.1
Накануне летального исхода нарастает
межгиппокампальная и полушарно-гиппокампальная когерентность ЭКоГ с максимумом
на частоте 4¬5 Гц (указано стрелкой)
3. В структуре межцентральных отношений
десинхронной составляющей на третьи сутки после деструкции ВЯД (рис. 4.4 НА)
отмечено резкое ослабление когерентных связей ствола с другими зонами мозга.
Тем не менее, на спектрах когерентности выделялся пик на частоте 6-8 Гц между
гиппокампом и корой -с неявным опережением по фазе ОФК, а также тенденция к
повышению внутриполушарного (лобно-сенсомоторного) взаимодействия в широком
частотном диапазоне - преимущественно справа. К пятым суткам, на фоне снижения
мощности высокочастотных составляющих ЭА, при сохранении на спектрограммах пика
лишь в дельта-диапазоне, наблюдалось дальнейшее падение связей как ствола с
другими образованиями, так и областей внутри полушарий (рис. 4.4 III A).
На спектрах когерентности синхронизированной
составляющей ЭА (рис. 4.4 II Б) на третьи сутки после операции связь между
стволом и другими зонами мозга сохранялась - однако, с формированием пиков на
других частотах: 4-5 Гц, 14 Гц и 22 Гц. Мало изменялись по сравнению с фоном
гиппокампально-корковые и корково-корковые связи, что указывало на их
определенную "прочность". Причем, структура этих связей не была
похожа на структуру стволовых и межполушарных когерентностей, что позволяет
думать об "автономном" характере внутриполушарной синхронизации.
Однако, к пятым суткам, т. е. накануне летального исхода (рис. 4.4 III
Б)
резко снизилась когерентность между стволом и другими зонами мозга (по всем
ритмам), симметричными лобными, а также лобной и сенсомоторной областями
(особенно в диапазоне от 0,5 до 12 Гц). При этом относительно усиливалась
межгиппокампальная (на частоте 4 Гц) и гиппокампально-корковая (на частотах
около 5 и 22 Гц) синхронизация. Таким образом, в условиях отчетливого
стволового угнетения и резкого снижения тонуса коры выявляются
электрографические признаки активации гиппокампальной синхронизирующей системы,
обеспечивающей, возможно, кратковременную стабилизацию гомеостаза.
Представленные результаты анализа ЭА у животных
с локальным стволовым повреждением демонстрируют значительное сходство динамики
ЭА крыс с изменениями ЭЭГ у больных с опухолевым стволовым повреждении в раннем
послеоперационном периоде. Сравнимы фазное течение с последовательным
повышением сочетанности определенных зон мозга (коры); различная частота
доминирующей синхронизации биопотенциалов тета-диапазона (6-7 или 4-5 Гц) при
разном характере послеоперационного течения (Гриндель с соавт., 1983; Шарова с
соавт., 1991, 1993). Это обусловливает важность представленных результатов
экспериментального исследования полушарно-гиппокампальных и
стволово-гиппокампальных отношений у животных, проведение которого затруднено у
человека.
Полученные данные позволяют ответить на ряд
вопросов, возникающих при трактовке послеоперационных ЭЭГ-изменений у человека.
1. Тождество
поведения тета-составляющих ЭА мозга крыс и ЭЭГ человека при остром стволовом
повреждении служит объективным доказательством единства этого вида активности и
подтверждает важную роль гиппокампа в его происхождении (Arnolds
et al.
1979; Bland, 1986; Haas,
1987).
2. При
отсутствии единого представления о механизмах генерации тета-ритма возможны
следующие предположения о нейрофизиологических механизмах формирования
разночастотной тета-активности, прогностически значимой для острых
патологических состояний человека: его частота может отражать как разное
функциональное состояние одной и той же функциональной системы, так и разную
организацию этой системы при разных типах стволового патологического очага
(стойкого или углубляющегося).
Эксперименты показали, что частотные параметры
синхронизации тета-активности (4-5 или 6-7 Гц) определяются, прежде всего,
состоянием стволово-гиппокампального входа, снижаясь по мере угнетения
функциональной активности ствола (Бражник, Виноградова, 1983) или
перераспределением ее в пределах стволовой медиаторной системы (Меликов 1987, Vertes
et al.,
1993). Полученные нами данные скорее подтверждают предположение о разном
состоянии одной и той же функциональной системы. Когерентный анализ ЭА позволил
как бы "визуализировать" особенности динамики
стволово-гиппокампального и гиппокампально-кортикального взаимодействия при
различных состояниях ретикуло-септального входа. Показано, например, что в
условиях относительной морфологической сохранности ствола именно этот вход
является "запускающим звеном" синхронизации на 6-7 герцах. Если же
состояние активирующей стволовой системы резко изменено (угнетено) вследствие
значительного объема стволового повреждения на уровне ВЯД (осложненное
течение), лидирующую роль в запуске этой реакции могут на некоторое время брать
корковые отделы мозга и гиппокамп. В наблюдениях с летальным исходом показана
возможность формирования автономной синхронизированной гиппокампально-
кортикальной системы на частоте 4-5 Гц, сопряженной с относительной
стабилизацией состояния низкоуровневого висцерального обеспечения.
3. У крыс подтверждено также участие ОФК в
формировании системной приспособительной реакции при остром стволовом
повреждении. Ранняя послеоперационная стволово-гиппокампальная
синхронизация
биопотенциалов
на частоте 6-7 Гц с последующим, а иногда и опережающим, участием в этой
реакции передних отделов коры характерны для выживших крыс. У погибшего
животного падение межполушарных связей ОФК предшествовало летальному исходу.
Следует отметить, однако, что у человека роль передних отделов полушарий в
послеоперационных адаптивных перестройках более существенна (Шарова с соавт.,
1993), что связано, по-видимому, с прогрессивным морфо-функциональным развитием
лобных отделов мозга в филогенезе.
Результаты экспериментальных исследований
свидетельствуют о том, что именно десинхронная составляющая ЭА мозга крыс
является "носителем" индивидуальных структурных реакций в процессе
послеоперационных системных перестроек ЦНС, а ритм 6-7 Гц - показателем
адаптивной стволово-гиппокампальной ирритации. Изменения же синхронизированной
составляющей ЭА, более выраженные при осложненном и летальном исходах, в
большей степени отражают, вероятно, состояние гомеостатического (висцерального)
управления. В этой связи обращает на себя внимание тот факт, что у одного из
погибших животных лишь накануне летального исхода отмечено снижение, помимо
стволовых, и межполушарных связей ОФК на синхронизированной составляющей ЭА,
что свидетельствует о причастности этих отделов мозга к процессам висцеральной
регуляции.
.1
Исследование роли гиппокампа в адаптивных реакциях цнс при остром стволовом
повреждении мозга крыс
Предыдущий раздел был посвящен разработке модели
локальной стволовой патологии, а также исследованию послеоперационных реакций,
развивающихся у животных при остром стволовом повреждении.
Установлено, что разные варианты послеоперационной
динамики состояния крыс сопровождаются характерными эмоциональными
проявлениями. Кроме того, была выявлена корреляция между уровнем
эмоциональности животных до операции, уровнем суммарной двигательной активности
(отражающих, по данным литературы, степень стрессоустойчивости крыс) и исходом
локального стволового повреждения (глава 3). Эти данные, наряду с результатами
наших электрофизиологических исследований (глава 4), подтверждают положения об
участии лимбической системы в формировании приспособительных реакций организма
на изменение внутренней и внешней среды. По данным литературы, это справедливо
и в отношении церебральных адаптивно-компенсаторных послеоперационных реакций
при остром повреждении ствола - как у человека (Шарова с соавт., 1991, 1992),
так и у животных (Gliddon
et al.,
2003).
Однако, специфическая роль отдельных образований
лимбической системы, в частности, гиппокампа и орбитофронтальной коры, в
приспособительных реакциях при экстремальных для организма ситуациях
исследована недостаточно. Для решения подобного рода проблем в физиологии
широко используются как электрофизиологические методы, так и методы разрушения
(или временного выключения) исследуемых структур. В нашей работе использованы
оба экспериментальных подхода: регистрация биоэлектрической активности (глава
4), а также повреждение методом электрической коагуляции определенных
лимбических образований. В рамках основной проблематики работы особый интерес
для нас представлял также анализ эмоциональных реакций оперированных животных.
Это обусловлено тем, что, по мнению ряда авторов, лимбическая система является
и главным регулятором мотивационно-эмоционального поведения млекопитающих
(Ведяев, 1974; Зилов, 1977; Ониани, 1980; Коваль с соавт.,1986; Шульговский,
1997; Судаков, 1998; Симонов, 2001).
Задачей настоящего раздела было проследить (по
широкому спектру поведенческих и клинико-неврологических показателей) динамику
функционального состояния и исходы после локального стволового повреждения
мозга крыс в сочетании с параллельным, двусторонним или односторонним (справа
или слева), нарушением целостности гиппокампа.
Известно, что гиппокамп имеет большое количество
афферентных входов, позволяющих получать обширную сенсорную информацию, а также
информацию от корковых и подкорковых структур (Гамбарян, Коваль, 1973; Отмахов,
1993). Важной афферентной системой гиппокампа является свод, куда поступает
основная масса сигналов, в том числе от гипоталамуса и ретикулярной формации
ствола мозга. Роль реле для этих восходящих путей выполняет перегородка. Если
сама перегородка играет ключевую роль в генерации тета-ритма гиппокампа
(Гусельников, 1976; Адрианов, 1999; Кичигина, Кудина, 2001), то его частота и
выраженность регулируется структурами ствола мозга через медиальный септум.
(Гасанов, Меликов, 1986; Heynen,
Bilkey, 1991; Виноградова
с соавт., 1995; Кичигина, Кудина, 2001).
Так как повреждение ствола мозга приводит к
нарушению механизмов саморегуляции и адаптационно-трофических влияний
регулирующих систем, таких как ретикулярная формация и лимбическая система
(Зимкина, 1958; Gliddon
et al.,
2003), очень важно тщательно исследовать участие гиппокампа, тесно связанного с
этими структурами, в приспособительных реакциях ЦНС при остром стволовом
повреждении. Нет полной ясности и в причастности гиппокампа к формированию тех
или иных эмоциональных реакций. С одной стороны, считается, что гиппокамп,
относясь к «информационным» структурам, принимает участие в формировании
эмоциональных реакций (Симонов,2001), что подтверждается и данными клиники (Брагина,
1974; Мадорский, 1985). Подтверждением этой точки зрения являются данные,
описывающие агрессивные реакции, сопровождающие повреждение гиппокампа -
подобные проявлениям стимуляции септума (Коваль с соавт., 1986), что, по мнению
авторов, может быть обусловлено тесным гиппокампально-септальным
взаимодействием. С другой стороны, ряд авторов утверждает, что у разных видов
животных повреждение гиппокампа зачастую не приводит к появлению отчетливых
изменений в мотивационно-эмоциональном поведении (Виноградова, 1975), что, как
считает Ониани (1980), может объясняться неполным (частичным) разрушением этой
структуры.
Всего было прооперировано 54 крысы. Но так как 8
крыс погибли в результате общего сепсиса, а две - после гиппокампального этапа
операции, то основу настоящего раздела нашей работы составили данные
динамического наблюдений 44 крыс с сочетанным стволово-гиппокампальным
повреждением головного мозга. Анализ данных до- и послеоперационного
исследования животных, с использованием статистических методов, позволил
выявить новые факты, способствующие более полному пониманию роли гиппокампа в
адаптивных реакциях при остром стволовом повреждении.
лимбический мозг гиппокамп
эмоциональный
Глава 5. Эффекты сочетанного стволового и
билатерального гиппокампального повреждения
Результаты, рассматриваемые в данной главе,
получены на 25 крысах, у которых электролитическая коагуляция ствола на уровне
латерального ВЯД сочеталось с аналогичным билатеральным повреждением СА1
гиппокампа (глава 2). При этом у 15 крыс сначала повреждали ствол мозга, через
1-1,5 суток проводили билатеральную электролитическую коагуляцию СА1
гиппокампа, а 10 крыс ствол мозга повреждался через 7 дней после билатерального
повреждения поля СА1. Из 25 прооперированных крыс 8 погибли в связи с наличием
общего сепсиса, связанного, вероятно, с общим снижением иммунитета. К этому
вопросу мы вернемся ниже. Две крысы погибли во время второй операции, связанной
с повреждением гиппокампа. В результате, группу с предшествующим стволовым
повреждением составили 9 животных. У 6 крыс вначале был поврежден гиппокамп.
Все выжившие прооперированные животные были забиты в период от 14 до 78 суток
после первого этапа операции (табл.5.1)
Каждую крысу до операции взвешивали, оценивали
эмоциональный фон, направление движения в открытом поле и характер моторной
межполушарной асимметрии. После операции ежедневно наблюдали за общим
состоянием оперированных крыс, определяя изменения в весе, характере
двигательной активности и эмоциональной сфере, состоянии кожного покрова и
шерсти, отмечая особенности развившихся неврологических нарушений (см. глава
2).
Послеоперационная динамика состояния
Как и при изолированном стволовом повреждении
(глава 3), все оперированные животные по характеру послеоперационного течения и
исходам были разделены на три группы: I- с неосложненным, II- с осложненным
послеоперационным течением и III - крысы с летальным исходом (табл.5.1).
Все крысы, у которых вначале повреждали ствол,
выжили. У двух особей в ходе послеоперационного наблюдения каких-либо неврологических
отклонений или существенных изменений в поведении не выявлено. По данным
гистологического исследования в одном из этих случаев имело место частичное
электролитическое повреждении стволовых ядер РФ в сочетании с односторонним
разрушением поля СА1 гиппокампа слева и очагом деструкции в корковой области
справа. В другом были слабо повреждены дорсальная часть кохлеарного ядра, а
также поля СА1 гиппокампа с двух сторон.
Таблица 5.1. Распределение по исходам животных
со стволово-гиппокампальным и изолированным повреждением ствола мозга.
Повреждение Исход
Ствол
+ гиппокамп
Ствол
+ гиппокамп
Гиппокамп
+ ствол
Всего
Ствол
Неосложненный
2
1
2(20%)
12(43%)
Осложненный
7
3
10(67%)
9(32%)
Летальный
0
2
2(13%)
7(25%)
Всего
9
6
15(100%)
28(100%)
Остальные семь животных после первого этапа
операции демонстрировали неврологическую симптоматику разной степени тяжести:
от наклона головы в оперированную сторону, не превышающего 30 градусов, до
грубых нарушений в виде винтообразных движений. Последующее повреждения
гиппокампа усугубляли эти нарушения. В двух случаях винтообразные движения
появились лишь после гиппокампального повреждения, имевшее место. Длительность
сроков уменьшения неврологического дефекта у этих 7 крыс колебалась от 11 до 78
суток. Эти дефекты не регрессировали полностью у четырех особей до момента
усыпления. Средние сроки редукции винтообразных движений при этом превосходят,
хотя и недостоверно, аналогичный показатель в группе с изолированной
коагуляцией ствола (см.табл.5.2 ).
К выявленным особенностям сочетанного
стволово-гиппокампального разрушения следует отнести также кратковременную (до
4 суток) потерю веса у части животных на
10-40 граммов с восстановлением его до исходного уровня в сроки от четырех до
тринадцати суток.
Таблица 5.2. Временные показатели
состояния животных при сочетанном стволово-гиппокампальном и изолированном
стволовом повреждении
Характеристики
Сроки
редукции после стволового этапа операции
Группы
Суммарная
стволовая симптоматика в случае неосложненного течения
Винтообразные
движения при осложненном течении
Кол-во
животных
Сроки
в сутках
Кол-во
животных
Сроки
в сутках
Ствол+гиппокамп
3
Симптомов
не было
6
1с.
7с. 7с** 9с. 30с. 36с
Ствол
4
2с.
2с. * 2с. 3.
5
3с.
3с.5с** 7с. 8с
Рис.5.1 Сопоставление сроков
редукции неврологических нарушений (А) и динамики эмоционального возбуждения
(Б) у крыс с сочетанным стволово-гиппокампальным повреждением со средним
дооперационным уровнем эмоциональности (группа с предшествующим стволовым
этапом операции). На рис.А - по оси абсцисс - сутки после стволового этапа
операции; по оси ординат-неврологические нарушения в баллах. 10 -винтообразные
движения; ноль соответствует норме; на рис.Б по оси ординат- формализованные
показатели эмоциональных реакций: 3 - агрессивно оборонительные; 2 -интенсивные
пассивно оборонительные; 1 - слабые пассивно оборонительные; 0 - отсутствие
эмоциональных реакций.
В данной группе оперированных крыс после второго
этапа операции (двустороннее повреждение гиппокампа) наблюдалось проявления
оборонительных эмоциональных (в том числе и агрессивно-оборонительных) реакций
с последующей их редукцией. Эти реакции сопровождали наиболее тяжелые
послеоперационные состояния.
Согласно данным гистологического исследования, у
5 животных из 7 разрушение разной глубины и объема на уровне ствола было
приурочено к структурам ВЯД, и у 2 - к области кохлеарного ядра. Что касается
области СА1 гиппокампа, то в 5 наблюдениях она была повреждена с двух сторон наряду
с коагуляцией вышележащих корковых областей; (см. рис.5.2); в двух остальных
случаях (№89, №90) имело место лишь изолированное корковое повреждение.
Рис. 5.2. Пример сочетанного
стволово-гиппокампального повреждения. Фронтальные срезы мозга крысы на уровне
поля СА1 гиппокампа (А) и ствола мозга (Б).
Окраска по методу Ниссля.
У 6 животных парное повреждение гиппокампа на 7
дней опережало стволовую коагуляцию. Две крысы из этой группы после второго
этапа операции погибли, остальные выжили.
Результатом первого - «гиппокампального» - этапа
операции было появление оборонительных реакций разной интенсивности, которые
исчезали к четвертым суткам. В этой группе животных следует отметить отсутствие
агрессивных оборонительных реакций, которые наблюдались у животных с
предшествующим повреждением ствола (см.рис 5.1) После второго этапа операции у
5 из 6 крыс, на фоне редукции эмоциональных реакций, выявились стволовые
неврологические нарушения (винтообразные движения, наклон головы под углом к
туловищу). Эти симптомы несколько регрессировали к 512 суткам, но сохранялись
до гибели (крысы №76, №77), либо усыпления (крысы №74, №78, №83) животных. По
данным гистологического исследования, попадание в ВЯД у 5 животных в двух случаях
сочеталось с двусторонним негрубым повреждением гиппокампа, а еще в трех - с
повреждением субикулюма.
Согласно данным патоморфологического
исследования погибших животных, у крысы №77, умершей на 18 сутки после
разрушения на уровне ствола и 25 сутки после повреждения гиппокампа было
выявлено выраженное полнокровие сосудов мягкой мозговой оболочки с
немногочисленными диапедезными кровоизлияниями (рис.5.3), что отражает
генерализованную неспецифическую реакцию сосудов на повреждение мозга. По
данным патоморфологических исследований, смерть наступила предположительно
вследствие начавшегося сепсиса.
У крысы №76, погибшей на 4-е сутки после
стволового этапа операции, обнаружен крупный абсцесс стволово-мозжечковой
локализации на фоне диффузного полнокровия сосудов. Однако, ни в том, ни в
другом случае в группе со стволово-гиппокампальной коагуляцией не отмечено
такой степени выраженности расстройств кровообращения, которая могла бы явиться
непосредственной причиной смерти животных. Наблюдаемые сосудистые реакции развивались
вторично по отношению к другим процессам, например, абсцессу (при более ранних
сроках смерти), или общих соматических расстройств (в более поздние сроки).
Здесь же уместно отметить, что у крыс №74 и №83 с осложненным течением
незадолго до усыпления мы наблюдали начальные проявления сепсиса (начальная
стадия фурункулеза). В трех случаях (крысы №77, №78, №83) были обнаружены
внутримозговые абсцессы в стволе или области корково-гиппокампального
разрушения на разных стадиях их формирования.
Рис.5.3 Гистологической препарат фронтального
среза больших полушарий мозга крысы №77 при стволово-гиппокампальном
повреждении с летальным исходом.
Увеличение х 100, окраска гематоксилином и
эозином.
Срез через орбитальную кору: полнокровие сосудов
мягкой мозговой оболочки (обозначено стрелкой). Массивные кровоизлияния
отсутствуют.
Суммируя экспериментальные данные, полученные в
группе животных с двусторонним повреждения гиппокампа можно заключить, что
стволово-гиппокампальная коагуляция, не увеличивая летальность по сравнению с
контрольной группой (с изолированным стволовым повреждением), приводит, главным
образом, к усилению выраженности у животных стволовой неврологической
симптоматики, а также к удлинению сроков компенсации нарушенных функций
(табл.5.2) Причем, когда коагуляция ствола опережала билатеральное повреждение
гиппокампа, стволовые симптомы могли появляться не сразу, а лишь после
гиппокампального этапа операции.
В целом, для всех случаев сочетанного
стволово-гиппокампального повреждения с выраженными стволовыми нарушениями, по
данным
гистологического
исследования, у животных имели место разрушения разного объема на уровне
ствола, приуроченные к структурам ВЯД (10 животных) и значительно реже (2
наблюдения) - кохлеарного ядра. Оно сочеталось с двусторонней коагуляцией поля
СА1 гиппокампа, зачастую вместе с вышележащими корковыми областями (8
наблюдений). Из 15 животных у трех не наблюдалось стволовой симптоматики. У
этих животных, по данным морфологического контроля, имело место либо негрубое
повреждение ядер стволовой ретикулярной формации, либо были незначительно
задеты кохлеарные ядра. Эти повреждения сочетались с двусторонней
субикулярно-гиппокампальной (СА1) деструкцией.
Вследствие сочетанного стволово-гиппокампального
повреждения погибли две крысы из 15, что составило 13% от общего числа
оперированных крыс. (Рис. 5.4). В то же время летальность животных в группе с
изолированной стволовой деструкцией составила 25% (глава 3). Следует отметить,
что при сочетанном повреждении ствола и гиппокампа летальный исход был
обусловлен преимущественно воспалительными процессами - как внутримозговыми
(абсцесс), так и системными (сепсис), в развитии которых определенную роль
могло сыграть (с учетом данных по этой группе в целом) резкое снижение общего
иммунитета, развившееся после нарушения целостности мозга.
Рис.5.4. Диаграмма распределения животных с
изолированным (А) и сочетанным (Б) стволово-гиппокампальным повреждением мозга
по исходам. 1 - осложненное течение; 2- летальный исход; 3 - неосложненное
течение;
Таким образом, особенность проявления
послеоперационных адаптивных реакций у животных со стоволо-гиппокампальным
повреждением состоит в нарастании стволовой симптоматики - особенно после разрушения
в гиппокампе. Она может быть объяснена данными литературы. Так, Русинова (1996)
показала, что электрическое раздражение гиппокампа вызывает усиление и
пролонгирование существования доминантных очагов (т.е. подкрепление),
сформированных в других отделах ЦНС - в частности, в стволе мозга. Косвенно это
подтверждается данными наших электрофизиологических исследований (глава 4) о
наличии в гиппокампе "резонансоподобных" электрографических реакций с
генерализацией на другие отделы мозга (у животных с неосложненным и осложненным
послеоперационным течением).
Обращает на себя внимание также высокий процент
животных со скрытыми или явными, локальными или диффузными воспалительными
процессами, что может быть связано с ослаблением иммунитета как следствия сочетанного
стволово-гиппокампального повреждения. Это утверждение согласуется с данными,
полученными другими авторами (Д.Ф. Плицитый,
С.В.
Магаева,
1970; Devi
et al.,1991,1993;
Pan et
al., 1993; Wetaiore
et al.,1994;
Магаева, Морозов, 2005). Так, в ряде работ 1990- 1993 годов Devi
et al.
указывают, что электролитическое повреждение или стимуляция гиппокампа
(вентрального) сказывается на состоянии иммунной системы. Разные области
гиппокампа могут либо стимулировать, либо подавлять гуморальную иммунную
систему (Pan
et al.,
1993). Считается (Wetmore
et al.,
1994; Магаева, Морозов, 2005), что гиппокамп, наряду с септумом и
гипоталамусом, включен в сложную и разнообразную регуляцию иммунной системы:
эти структуры могут избирательно участвовать в регуляции нейроиммунных ответов
и оказывать облегчающий эффект на иммунный ответ у самок крыс. Плицитый и
Магаева (1970) утверждают, что гиппокамп оказывает регулирующее влияние на
иммуногенез.
Предполагаемое нами наличие сдвигов в иммунной
системе крыс с сочетанным
стволово-гиппокампальным повреждением (не характерное для локального стволового
патологического очага) не противоречит данным морфологии.
Известно (Ониани,1980), что волокна повреждаемой нами области гиппокампа входят
в систему дорсального форникса, которая, как отмечено в главе 2, обеспечивает
связи гиппокампа с мезодиэнцефальными структурами. Эти взаимодействия
обусловливает роль гиппокампа в качестве модулятора эмоциональных,
биоэлектрических, вегетативных реакций (Болдырева с соавт., 1972; Брагина,
1974; Хамильтон, 1984; Симонов, 1993) и участие в регуляции работы иммунной
системы.
Изменения в эмоциональной сфере
Наряду с основной исследуемой проблематикой мы
рассматривали также вопрос о том,
влияет ли (и каким образом) сочетанное повреждение
ствола
и билатерального гиппокампа на эмоциональную сферу животных. Для
этого
с помощью теста Кинга, проведенного до операции, оценивали такие показатели
как: реакция на касание к спине и к носу, мышечный тонус при
взятии
на руки, наличие уринаций и дефекаций, а также вокализации (см. главу 2). По
совокупности признаков все животные делились на спокойных и
эмоциональных.
Рис.5.5 - внутренний круг - соотношение
спокойных и эмоциональных крыс до операции, наружный круг - после операции).
1
- спокойные животные; 2 - эмоциональные животные; А - сочетанное
стволово-гиппокампальное
повреждение; Б - изолированное стволовое повреждение.
Сопоставлялась доля выраженности тех и других (в
%) до и после операции - с оценкой значимости различия в этих соотношениях по
критерию хи-квадрат. Для наглядности эти данные представлены в виде круговой
диаграммы на рис.5.5 А.
После билатерального повреждении гиппокампа в
сочетании со стволовым у некоторых животных наблюдались бурные эмоциональные
реакции - вплоть до агрессивно-оборонительных. В целом по группе увеличивалась
доля эмоциональных крыс по сравнению с дооперационным уровнем на 14%: до
операции было 43% спокойных и 57% эмоциональных; после операции 29% спокойных и
71% эмоциональных. Однако, по критерию хи-квадрат = 2.25 эти изменения
соотношений недостоверны. Такой результат может быть обусловлен как
недостаточным числом наблюдений, так и разным их количеством до и после
операции. Объяснить появление эмоциональных реакций после повреждения поля СА1
гиппокампа можно если вспомнить, что после разрушения поля СА1 наиболее сильно
страдают дифференцировочное и угасательное торможение (Валюх, 1981; Карамян,
Соллертинская, 1982). Поэтому ряд авторов полагает, что гиппокамп принимает
участие в регуляции тормозных процессов, обеспечивая отбор необходимой
информации из хранилищ памяти. И его повреждение может приводить к появлению
«биологически отрицательных реакций» (Урманчеева, 1970; Карамян, Соллертинская,
1976; Лагутина с соавт., 1976; Коваль, Саркисов, 1976; Адрианов, 1999). Правда,
данные Левшиной с соавт. (1977), об изменении поведения крыс в сторону
преобладания процессов торможения (уменьшение двигательной активности,
агрессивности, выраженная сомноленция) после двустороннего повреждения
гиппокампа, противоречит нашим данным. Возможно, объяснением этого противоречия
является сочетание гиппокампального повреждения со стволовым в нашем случае, а
«биологически отрицательные реакции» обусловлены позными нарушениями, что можно
отнести к «трудному состоянию» (Коваль, Саркисов, 1976).
Для сравнения были проведены сопоставления по
группе животных с изолированным стволовым поражением (рис. 5.5). Выявленные
аналогичные изменения соотношений разно эмоциональных крыс, также статистически
недостоверны: 62% спокойных и 38% эмоциональных до операции; 48% спокойных и
52% эмоциональных после операции (т.е. увеличение «эмоциональных» крыс в группе
на 14% недостоверно).
Следует отметить, что при изолированном
стволовом повреждении агрессивно-оборонительные реакции имели место только
перед летальным исходом. В остальных случаях эмоциональные реакции носили
пассивно-оборонительный характер. При сочетанном стволово-гиппокампальном
повреждении, как уже отмечалось, в том случае, когда сначала повреждали ствол,
у животных с осложненным характером течения послеоперационного периода
наблюдались агрессивно-оборонительные реакции. По мере восстановления
эмоциональные реакции исчезали. Если повреждение гиппокампа производилось
сначала, то эмоциональные реакции также исчезали по мере восстановления позы и
движения. Но в этом случае следует отметить отсутствие
агрессивно-оборонительных реакций. Каких-либо качественных отличий в протекании
эмоциональных реакций разного знака у животных с сочетанным
стволово-гиппокампальным повреждением нами обнаружено не было.
Таким образом, сочетанное повреждение ствола и
симметричных отделов гиппокампа приводит к увеличению в выборке доли
эмоционально реактивных животных. В этом случае эмоциональные реакции более
выражены, чем при изолированном повреждении ствола. Однако, эти изменения не
являются статистически достоверными.
Глава 6. Особенности латерализованного
повреждения гиппокампа на фоне острого стволового патологического
очага
При изучении роли гиппокампа в
адаптивно-компенсаторных процессах головного мозга нельзя не учитывать того
обстоятельства, что данная структура является парной, билатерально
симметричной, соединенной между собой комиссурами (Гамбарян, Коваль, 1973;
Ониани, 1980; Хамильтон, 1984; Van
Groen, Wyss,
1988; Отмахов, 1993).
Определенная независимость в деятельности обоих
гиппокампов была обнаружена М. Брейже еще в
1967 г. К настоящему времени накоплены убедительные доказательства того, что
гиппокампальные образования правого и левого полушарий могут действовать как
две независимые субъединицы. Экспериментальные исследования И.П.
Левшиной
с соавт. (1977) свидетельствуют о неоднозначной роли правого и левого
гиппокампа в процессах обучения, поддержания эмоционального напряжения и
регуляции вегетативных функций. Авторы пришли к выводу о биохимической природе
латерализации функций гиппокампа. Относительная функциональная независимость
симметричных отделов гиппокампа была показана и в других работах (Chida,
Toyosawa, 1991; Maguire
et al.,
1997, 2003; Beauregard
et al.,
1998; Молодцова, 1999; Артюхина, Саркисова, 2000; Kristofikova
et al.,
2004). Особенно следует подчеркнуть, что асимметрия гиппокампа была
подтверждена электрофизиологическими исследованиями (Квирквелия, 1987; Chida,
Toyosawa, 1991). Однако,
как справедливо отмечает П.В. Симонов
(1999), данные о функциональной асимметрии гиппокампа, в основном, получены на
человеке, в условиях клиники (Брейже, 1967; Брагина, 1966, 1967; Gray,
1972; Мадорский, 1985; Beauregard,
1998; Болдырева, 2000а,б). Как считает Н.Н. Брагина на основании только
клинических данных неправомочно говорить о функции собственно гиппокампа
(Брагина, 1966, 1967). Поэтому особенно важно представляется уточнение роли
симметричных областей гиппокампа в адаптивно-компенсаторных перестройках мозга
в условиях эксперимента.
В настоящей работе, наряду с моделями
сочетанного стволового и двустороннего повреждения гиппокампа, мы рассматривали
ситуацию повреждения ствола в сочетании с односторонним повреждением
гиппокампа. Экспериментальную группу составили 19 животных: 15 - самцов
беспородных белых крыс, 4 - самца линии Wistar.
Последние существенно отличались от основной группы по возрасту и
использовались нами лишь как индивидуальные наблюдения, но были исключены из
общего статистического анализа (Мельников с соавт., 2001). У всех животных (за
исключением крысы № 45) за 7 дней до операции на стволе мозга производили
одностороннее повреждение гиппокампа: у 8 из 15 беспородных крыс с левой
стороны, у 7 - с правой.
Следует отметить, что в группе из 19
отсутствовали летальные послеоперационные исходы.
Эффекты сочетанного повреждения ствола и левого
гиппокампа Послеоперационная динамика состояния
У двух крыс (№86, №101) с сочетанным
повреждением ствола и левостороннего гиппокампа явных признаков патологии после
операции выявлено не было. Они были отнесены к варианту легкого
послеоперационного течения (глава 3). Морфологический контроль у этих животных
выявил наличие умеренных повреждений поля СА1 гиппокампа слева в сочетании с
зонами деструкции на уровне ствола. Последние располагались существенно
латеральнее и выше ВЯД: в tuberculum
acusticum, кохлеарных ядрах
и у основания ножки мозжечка.
Большая часть животных этой группы (6 из 8, т.е.
75% наблюдений) по особенностям послеоперационной динамики было отнесено к
осложненному варианту (глава 3). У четырех крыс имели место винтообразные
движения, исчезающие на 5-9-е сутки после повреждения ствола мозга. Исключение
составляла лишь крыса №93, у которой винтообразные движения исчезли только на
36-е сутки после стволового этапа операции. У этих животных имело место
длительное сохранение менее грубых неврологических отклонений, таких как наклон
головы в оперированную сторону. Еще у одной крыс (№102) наблюдался хотя и
слабый, но длительно сохраняющийся (33 дня) наклон головы. Следует отметить, что
у особей данной группы отмечено существенное падение веса в первую 1-1,5 недели
после стволового этапа операции, зачастую большее, чем при двустороннем
повреждении гиппокампа. Потери веса могли достигать 40-60 граммов и имели место
у животных с наличием винтообразных движений (крысы №92, №93, №96, №98). Через
10-12 суток вес восстанавливался.
Морфологические исследования показали, что в
большинстве случаев у крыс с осложненным течением послеоперационного периода
имело место повреждение поля СА1 гиппокампа в сочетании с полным левосторонним
разрушением ВЯД, за исключением двух животных. У крысы №98 латеральная часть
ВЯД была лишь задета. В основном, разрушение было локализовано в латеральном
кохлеарном ядре. В другом наблюдении (крыса №102) ядро Дейтерса практически не
было задето, но основательно разрушена область fibrae
vestibulocerebelares.
Представляет интерес течение послеоперационного
периода у крысы №45. Исходно она относилась к группе с двухсторонним поражением
гиппокампа, однако морфологический контроль выявил повреждение этой структуры
только слева. Поэтому данное наблюдение расценивалось как пример сочетанного
стволово-левогиппокампального повреждения, когда стволовый этап операции
предшествовал гиппокампальному.
У этой крысы в первые сутки после коагуляции
ствола мозга отмечались лишь слабые динамические нарушения позы: при движении
на плоскости передняя часть туловища была повернута под углом к задней половине
тела, хотя винтообразные движения отсутствовали. К 3-4 суткам после стволового
и 2-3-м после гиппокампального этапа операции наблюдалось постепенное
усугубление
тяжести общего состояния животного. Это выражалось в ухудшении состояния шерсти
и существенной потере веса (с 210 до 160 гр.). На 5 - 7 сутки у крысы
выявлялись также винтообразные движения. После 10-х суток животное стало
набирать вес и его состояние стало улучшаться. Поза восстановилась через месяц.
Исходно спокойная, эта крыса на протяжении всего послеоперационного периода
(вплоть до окончательного восстановления позы) отличалась повышенной
эмоциональностью.
Рис. 6.1. Сравнение сроков редукции
неврологических отклонений (А) и уменьшение уровня эмоционального возбуждения
(Б) у крысы №45 с сочетанным стволово-левогиппокампальным повреждением. По оси
абсцисс в обоих случаях - сутки после операции. По оси ординат: (А) -
ранжированные показатели неврологических отклонений. Максимальные отклонения в
виде винтообразных движений соответствуют 10 баллам. (Б) - формализованные
показатели эмоциональных проявлений (1 - слабая оборонительная реакция; 2 -
сильная оборонительная реакция; 3 - агрессивно-оборонительная реакция.
Следует отметить увеличение уровня
эмоциональности при ухудшении общего состояния, а также усугубление позных
нарушений после левостороннего повреждения гиппокампа. Появление винтообразных
движений в данном случае может быть объяснено "наложением" эффектов
повреждения ствола и одностороннего повреждения гиппокампа.
Сходные данные получены в работе Н.И.
Артюхиной
и К.Ю.
Саркисовой
(2000, 2004): двусторонняя окклюзия сонных артерий сопровождалось развитием у
животных судорожных припадков (с последующим летальным исходом) и появлением у
выживших животных вращательных движений.
Морфологический контроль выявил у этих животных
очаги ишемического инсульта, локализовавшегося преимущественно в СА1 правого
гиппокампа - с исчезновением в зоне инсульта нервных клеток и пролиферацией
глии с образованием глиального рубца. Появление такого рубца и было, вероятно,
причиной судорожных припадков. По данным клиники, окклюзия сонных артерий зачастую
сопровождается отеком мозговой ткани с вторичной дисфункцией ствола (Саркисова,
2000). Вовлечение в патологический процесс стволовых образований (наряду с
повреждением гиппокампа) и обусловливало, по-видимому, возникновение устойчивых
нарушений в виде винтообразных движений.
Изменения в эмоциональной сфере
У крыс этой группы оценивали также соотношение
«спокойных» и «эмоциональных» животных до и после операции (рис.6.3).
Рис.6.2. Диаграмма, отражающая эмоциональность
животных до операции (внутренняя окружность) и после (наружная окружность)
сочетанного повреждения ствола и гиппокампа слева (А) или ствола и гиппокампа
справа
(Б).
1
- спокойные животные; 2 - эмоциональные животные.
Представленная диаграмма демонстрирует, что
сочетанное повреждение ствола и поля СА1 гиппокампа слева вызывает увеличение
количества «эмоциональных» животных с 22% до 78% (т.е. на 56%), что является
статистически значимым по критерию хи-квадрат ( %2=4; р > 3.84).
Следует отметить отсутствие агрессивных реакций
у крыс этой группы. Наблюдаемые эмоциональные реакции даже небольшой
интенсивности сохранялись длительное время, иногда (крысы №98 и №102) до
окончания наблюдений. При этом интенсивность эмоциональных реакций не
коррелировала с тяжестью исхода. Эффекты сочетанного повреждения ствола мозга и
правого гиппокампа
Послеоперационная динамика состояния
У одной из 6 крыс (№99) послеоперационное
течение расценивалось как легкое: после стволового этапа операции у нее
выявлялся слабый наклон головы, который редуцировался к третьим суткам. По
данным морфоконтроля, при наличии повреждения поля СА1 правого гиппокампа, в
стволе область разрушения располагалась латеральнее ВЯД, и можно предположить,
что позные нарушения возникли в результате электротонических влияний.
У 5 из 6 крыс (т.е. почти в 83%) позные
нарушения были более тяжелыми. У 3 крыс (№91, №103, №104)отмечались
винтообразные движения, исчезавшие на 2-9 сутки после стволового этапа
операции. Лишь в одном случае (крыса №105) "винт" сохранялся до 12
суток. При этом у крыс с грубыми нарушениями процесс восстановления отличался
от такового у животных с левосторонним повреждением гиппокампа в сочетании со
стволовым. Отличие заключалось в большем количестве переходных состоянием и
более длительном сохранении их во времени. Так, при левостороннем повреждении
гиппокампа сразу после исчезновения винтообразных движений наблюдались наклон
туловища и наклон головы свыше 60 градусов, которые сохранялись иногда до трех
месяцев. При правостороннем повреждении гиппокампа длительное время сохранялась
неспособность крыс удерживаться на ногах: животные лежали на оперированном боку
до 20-75 дней.
Морфоконтроль мозга животных этой выявил наличие
деструкции в поле СА1 у 5 животных. При этом у трех крыс (№91, № 104, №105)
имело место полное разрушение ВЯД. У двух животных (№97, №103) ВЯД было задето
в верхней части, а основные разрушения относились к ножке мозжечка. У крысы №99
разрушения ВЯД, как уже было сказано выше, не обнаружено. В целом характер
разрушений сопоставим с таковым в предыдущей группе животных. Тяжесть
неврологических отклонений пропорциональна величине повреждения Изменения в
эмоциональной сфере
Сопоставление эмоциональности животных до и
после сочетанного повреждения ствола
и правого гиппокампа продемонстрировало самое большое
(на
60%) и статистически достоверное по критерию хи -квадрат (X2 = 5; р
> 3.84) увеличение
"эмоциональных" животных после операции среди всех
рассматриваемых
в разделе групп наблюдений (рис. 6.3).
Рис.6.3. Диаграмма, отражающая изменение
количества эмоциональных животных после сочетанного повреждения поля СА1
гиппокампа с правой стороны и ствола мозга. Внутренняя окружность - соотношение
спокойных и эмоциональных животных до операции; наружная - то же самое после
операции. 1 - спокойные; 2 -
эмоциональные
Сравнение двух групп показало, что у крыс с
правосторонним повреждением гиппокампа эмоциональные реакции были более
интенсивные, чем в группе с левосторонним повреждением. Но интенсивность
реакций не была постоянной, а
носила ярко выраженный волнообразный характер. Не удалось обнаружить связь
между появлением или усугублением эмоциональных реакций и изменениями
неврологического статуса, как это имело место при изолированном повреждении ствола
или при сочетании двустороннего повреждения гиппокампа со стволом мозга.
Следует также отметить, что при увеличении общего количества эмоциональных
животных наблюдалось отсутствие агрессивных реакций при одностороннем
повреждении гиппокампа (независимо от стороны повреждения) в сочетании со
стволом.
Таким образом, в результате проведенных
исследований нам удалось уточнить значимость гиппокампа в церебральных
приспособительных реакциях при остром повреждении ствола мозга крыс. Показано,
что сочетанная стволово-гиппокампальная коагуляция усугубляет, по большей
части, стволовую симптоматику и пролонгирует время ее обратного развития, т.е.
вызывает подкрепление очага стойкой патологической активности стволовой
локализации. Повреждение гиппокампа в сочетании со стволовым приводит к
нарушению иммунитета у крыс.
При латерализованном повреждении гиппокампа в
сочетании со стволом следует отметить отсутствия смертельных исходов и
нарушений в деятельности иммунной системы. Сочетанное повреждение ствола с
полем СА1 правого или левого гиппокампа вызывает достоверное увеличение
эмоциональности большинства крыс, в отличие от крыс с билатеральным
повреждением гиппокампа.
Установлено также, что правый и левый гиппокамп
вносят свои качественные особенности в компенсаторный процесс при осложненном
течении послеоперационного периода. Так, при правостороннем повреждении
послеоперационное восстановление шло медленнее и сопровождалось длительным
сохранением грубых нарушений (неспособность удерживаться на лапах). При
левостороннем повреждении грубые нарушения (винтообразные движения) исчезали на
2-9-е сутки после операции. Однако нарушения позы в виде наклона головы в
оперированную сторону сохранялись длительное время.
6.1
Исследование роли орбитофронтальной коры в адаптивных реакциях цнс при остром
стволовом повреждении мозга крыс
Данные нейрофизиологических исследований у
больных с очаговым поражением ствола (Шарова с соавт. 1993, 1995), а также
собственные исследования биоэлектрической активности мозга крыс после
повреждения ствола на уровне ядра Дейтерса (глава 4) свидетельствуют о
возможном включении в послеоперационные адаптивно-компенсаторные процессы,
наряду с гиппокампом, лобных отделов больших полушарий мозга.
Целью настоящего раздела работы был анализ
динамики функционального состояния и послеоперационных исходов у 33 животных с
локальным стволовым повреждением мозга в сочетании с двусторонним, либо
односторонним (справа или слева) нарушением целостности орбитофронтальной коры
(ОФК).
Как отмечалось ранее (см. гл.2), выбранная нами
область коагуляции ОФК является проекционной зоной медиодорсального ядра
таламуса. Эта область рассматривается рядом авторов как лимбическая кора и
является аналогом средних отделов поля 24 (Василевская, 1971; Замбржицкий,
1972; Divac, 1972;
Беллер, 1977; Пигарева, 1984; Хамильтон, 1984).
Глава 7. Эффекты сочетанного стволового и
билатерального
орбитофронтального повреждения
Группу с сочетанным электролитическим
повреждением ствола и билатеральным повреждением ОФК составили 16 животных.
У 12 из них разрушение ствола на 1-2 суток
опережало двустороннюю электролитическую коагуляцию ОФК; у остальных особей
парное повреждение ОФК на 7 дней опережало повреждение ствола мозга. Все
выжившие животные были забиты в период от 14 до 78 суток после первого этапа
операции для последующего морфоконтроля. Послеоперационная динамика состояния
По аналогии с предыдущими исследованиями,
животные с сочетанным стволово-орбитофронтальным повреждением были разделены на
три группы по характеру послеоперационного течения и исходам: I - с
неосложненным течением, II - с осложненным течением и III - крысы с летальным
исходом (табл. 7.1).
Таблица 7.1 Сравнение
распределения по исходам животных с разным типом повреждений (ствол в сочетании
с билатеральным повреждением ОФК, а также изолированное повреждение ствола
мозга)
Повреждение Исход
Ствол
+ ОФК
Ствол+ОФК
ОФК
+ ствол
Всего
Ствол
Неусложненный
5
0
5(31%)
12(43%)
Осложненный
0
1
1(6%)
9(32%)
Летальный
7
3
10(63%)
7(25%)
Всего:
12
4
16(100%)
28(100%)
Полученные нами данные свидетельствуют о разных
последствиях изолированного, а также сочетанного стволово-орбитофронтального и
стволово-гиппокампального повреждений.
Как видно из предыдущего материала (раздел II),
при стволово-гиппокампальном повреждении наблюдается нарастание тяжести
состояния животных по сравнению с изолированной деструкцией на уровне ствола.
При этом у крыс усугубляется стволовая неврологическая симптоматика и
увеличивается время ее обратного развития. Сочетанное же
стволово-орбитофронтальное разрушение с вовлечением латерального вестибулярного
ядра Дейтерса приводит к более тяжелым результатам - независимо от
последовательности воздействия (таблица 7.1; рис.7.1). Из 16
экспериментальных
крыс этой серии погибли 10 животных, что составляет 62,5 процента и значимо (по
критерию хи-квадрат) превышает летальность как в группе с изолированным
стволовым (%2= 7.89; р= 0.005), так и с сочетанным
стволово-гиппокампальным поражением.
Рис.7.1 Диаграмма распределения животных с изолированным
стволовым (А) и сочетанным стволово-орбитофронтальным (Б) повреждением мозга по
исходам.
1-
осложненное течение послеоперационного периода; 2 - летальный исход; 3 -
неосложненное течение послеоперационного периода.
В первую очередь мы рассмотрим животных с
летальным исходом. У 7 из 10 крыс этой группы повреждение ствола предшествовало
повреждению ОФК на 1-2 суток. Эти крысы погибали на первые - восемнадцатые
сутки после стволового этапа операции.
Неврологические нарушения у двух животных (№31,
№54) после повреждения ствола были умеренными (изменение тонуса передних и
задних конечностей), а еще у пяти крыс (№№ 36, 40, 41,56, 62) - грубыми. Эти
крысы потеряли способность передвигаться на плоскости, у них появились
винтообразные движения.
По сравнению с изолированным повреждением ствола
мозга разрушение ОФК привносило в клиническую картину ряд особенностей:
усугубление неврологического дефекта на фоне проградиентного нарастания тяжести
состояния в виде значительной потери веса (от 18 до 50%), иногда стремительной,
что, возможно, свидетельствует о вторичном вовлечении в патологический процесс
гипоталамических областей. Кроме того, имели место трофические нарушения в
форме сукровичных подтеков вокруг глаз и некротических пятен около рта,
загрязнение шерсти. В некоторых наблюдениях отмечено нарушение питьевого
поведения (повышенная жажда либо, напротив, отказ от воды) - также,
по-видимому, вторично-гипоталамического характера. В отдельных случаях выявлены
отчетливые изменения глубины и ритма дыхания.
У трех животных (№№70, 71, 73) группы с
летальным исходом повреждение орбитофронтальной коры на 7 дней опережало
коагуляцию ствола. Эти крысы погибли на 1, 13 и 18 сутки после стволового этапа
операции. Послеоперационная динамика их состояния была сходной с динамикой
состояния у погибших животных из предыдущей серии. Рис.7.2 Пример сочетанного
стволово-орбитофронтального разрушения. Фронтальные срезы мозга крысы на уровне
передних отделов коры (А) и ствола мозга (Б). Стрелками указаны области
разрушения. Окраска по методу Ниссля.
Гистологическое исследование мозга крыс с
опережающим стволовым повреждением (проведенное у 6 из 7 животных) выявило
наличие обширного левостороннего электролитического разрушения на уровне
ствола: в четырех случаях в ядре Дейтерса, в одном - в кохлеарном ядре. У 5
крыс при этом имело место двустороннее разрушение ОФК (рис.7.2). В одном случае
оно было лишь правосторонним (крыса №62).
У крыс с опережающим повреждением
орбитофронтальной коры, по данным гистологического исследования мозга,
левостороннее электролитическое разрушение ядра Дейтерса сочеталось с
двусторонней, либо обширной левосторонней (№71) коагуляцией орбитофронтальной
коры.
Патоморфологическое исследование погибших
животных показало, что причиной летального исхода во всех случаях явилось
острое расстройство кровообращения по геморрагическому типу. Оно проявлялось в
резчайшем полнокровии сосудов мозга и оболочек, субарахноидальных
кровоизлияниях, отеке мозга, в одном наблюдении (№41) - в массивных
внутримозговых геморрагиях с прорывом в желудочки мозга; еще в одном случае
вышеописанные изменения сопровождались гемотампонадой всей желудочковой системы
(рис. 7.3)
Рис. 7.3. Гистологический препарат фронтального
среза больших полушарий мозга крысы при стволово-орбитофронтальном повреждении
с летальным исходом
Срез сделан на уровне боковых желудочков. Имеет
место гемотампонада желудочков (указано
стрелками).
Увеличение х100, окраска гематоксилином и
эозином.
Особенно следует подчеркнуть, что описанные
расстройства кровообращения носили острый диффузный характер и распространялись
на все отделы мозга: большие полушария, стволовые отделы, мозжечок.
В группе с летальным исходом при опережающем
повреждении орбитофронтальной коры по отношению к стволу выявленные нарушения
кровообращения характеризовались лишь несколько меньшей интенсивностью.
Группу выживших животных составляли 6 крыс. У
пяти из них коагуляция ствола предшествовала повреждению ОФК. При отсутствии изменений
в неврологическом статусе после первого и второго этапов операции (крысы №№55,
57, 58) морфоконтроль мозга этих крыс выявил стволовую коагуляцию кохлеарного
ядра или ножек мозжечка без повреждения ядра Дейтерса, сочетавшееся с
билатеральным (2 наблюдения) либо левосторонним (1 животное) повреждением
орбитофронтальной коры.
У двух выживших крыс (№№35, 63) после операции
на стволе мозга отмечались негрубые, но достоверно более стойкие, чем у
животных с изолированным повреждением ствола неврологические симптомы в виде
наклона головы влево и изменение тонуса задних конечностей - без изменения
структуры неврологического дефекта после орбитофронтального этапа операции
(табл. 7.2)
Таблица 7.2. Временные показатели состояния
животных при сочетанном стволово-орбитофронтальном и изолированном стволовом
повреждении.
Характеристики групп
Суммарных
стволовых симптомов при неосложненном течении
Винтообразных
движений при осложненном течении
Кол-во
животных
Сроки
в сутках
Кол-во
животных
Сроки
в сутках
Ствол
+ ОФК
2
*14с;
21с
1
35
**
Ствол
4
*2с;
2с; 2с; 3с
5
**3с;
3с; 5с; 7с; 8с
* Различие средних (по критерию Стьюдента)
статистически достоверно р < 0.002; ** р < 0.17
В одном из этих наблюдениях стволовой
коагулирующий электрод повредил ножку мозжечка (№35), в другом (№63)-
кохлеарное ядро слева. Разрушение ОФК у крысы №63 было левосторонним.
В том случае, когда повреждение
орбитофронтальной коры на 7 дней опережало коагуляцию ядра Дейтерса, выжила
одна крыса. Она была отнесена в группу с осложненным послеоперационным течением
(см. табл. 7.1), т.к. у нее сохранились рецидивы винтообразных движений в
течение 35 суток, а неспособность управлять задними конечностями сохранялась до
четырех месяцев после операции.
Гистологическое исследование мозга выявило в этом
случае негрубую коагуляцию ядра Дейтерса и незначительное двустороннее
повреждение орбитофронтальной коры.
Стволово-орбитофронтальные разрушения, как уже
было сказано ранее, помимо усугубления стволовой неврологической симптоматики,
вызывали значительное нарастание тяжести общего состояния, выраженные
трофические нарушения и потерю веса (рис. 7.3). Рисунок 7.3 иллюстрирует
изменение веса во всех группах экспериментальных животных: как со
стволово-орбитофронтальными, так и со стволово-гиппокампальными разрушениями -
при разных исходах. На графике видно, что вес у животных со
стволово-гиппокампальном повреждением больше, чем у крыс со
стволово-орбитофронтальными разрушениями. Различия веса недостоверны (на уровне
значимости 5%), что обусловлено малым количеством животных в сопоставляемых
группах. Тем не менее, следует отметить факт систематического превышения веса у
животных со стволово-гиппокампальными повреждениями по сравнению со
стволово-орбитофронтальными.
Рис.7.4. Изменения веса в группах
крыс с неосложненным (А) и осложненным (Б) послеоперационным течением, а также
летальным исходом (В) при сочетанном стволово-гиппокампальном (сплошная линия)
и стволово-орбитофронтальном (пунктирная линия) повреждении мозга.
По оси абсцисс - сроки после
стволового этапа операции: 1 - 1-3 сутки; 2 - 4-8 сутки; 3 - 9-15 сутки; 4 -
16-20 сутки; 5 - 21-30 сутки; 6 - 31-50 сутки; 7 - 51-80 сутки.
По оси ординат - усредненные по
группам наблюдений отклонения веса (в граммах) относительно дооперационного
уровня.
При этом на некоторых стадиях вероятность ошибки
относительно различий средних по критерию Стьюдента ниже 0.2 или 0.08, что
позволяет предполагать существование
локализационно-специфических
послеоперационных
изменений веса: при стволово-орбитофронтальном повреждении имеет место большее
его падение. Следует отметить,
что у других исследователей потеря веса имела место при двусторонней
экстирпации заднего отдела поясной извилины. Так И.А.
Замбржицкий
(1972), ссылаясь на работы В.М. Касьянова
(1947, 1949), отмечает потерю веса у собак при экстирпации именно заднего
отдела поясной извилины. Экстирпация же переднего отдела приводит, наряду с
другими вегетативными расстройствами, к длительным колебаниям диуреза и
нарушениям в нейрорегуляции колебания сахара в крови (Замбржицкий, 1972). Так
как явления диуреза наблюдались и у наших экспериментальных животных, то
возможно, одной из причин колебания веса в нашем случае было нарушение
деятельности желудочно-кишечного тракта. Второй причиной, как уже было сказано
выше, может быть вовлечение в процесс гипоталамических структур.
Анализируя полученные данные в контексте
проблемы специфической значимости разных отделов лимбической системы в
висцеральной регуляции, можно говорить о причастности орбитофронтальной коры,
например, к формированию системных сосудистых реакций (Замбржицкий, 1972;
Беллер, 1977; Шмидт, Тевс, 1985). Роль ОФК, как оказалось, приобретает особо
важное значение при повреждении ствола, в котором расположены основные центры
иерархически организованной системы сосудистой регуляции (Шмидт, Тевс, 1985) -
особенно на ранних (до двух недель) послеоперационных этапах, когда включение
этого палеокортикального "функционального дублера" становится
жизненно необходимым. У экспериментальных животных нарушения кровообращения
могут быть обусловлены, в частности, тем обстоятельством, что в лобных долях
находятся проекции норадренергической медиаторной системы (Harley,
1987), причастной к регуляции сосудистого тонуса (Amaral,
Simmanov, 1977). Результаты
исследования свидетельствуют о целесообразности дифференцированного изучения
участия правой и левой орбитофронтальной коры в регуляции висцеральных функций.
Кроме того, можно сказать, что некоторые соматические послеоперационные
проявления в этой группе животных (нарушение пищевого поведения и трофики,
потеря веса) свидетельствуют также о развитии вторичной послеоперационной
дисфункции гипоталамуса.
Изменения в эмоциональной сфере
В литературе имеются многочисленные данные о
непосредственном участии передних отделов новой коры в организации мотиваций и
эмоций (Замбржицкий, 1972; Хананашвили, 1972, 1983; Симонов, 1993; Adolphs,
2002; Cardinal
et al.,
2002; Davidson,
2002). Так, Наута (1963) рассматривал лобную область как неокортикальное
продолжение лимбической системы. Участие в формировании эмоциональных реакций
передних отделов мозга отмечал и ряд других авторов. Как было сказано ранее, по
данным Замбржицкого двусторонняя цингулоэктомия у животных сопровождалась
реакцией со стороны внутренних органов. Наблюдалось усиление моторики
желудочно-кишечного тракта, расширение зрачков, гиперсекреция слюны и др. Имела
место гиперактивация блуждающего нерва, парасимпатических веточек
глазодвигательного и лицевого нервов. Автор пришел к выводу, что эти проявления
связаны с нарушением синергии парасимпатической и симпатической регуляции.
Наряду с этими последствиями при разрушении или
стимуляции лимбических отделов коры у разных видов животных наиболее часто
нарушается сердечная деятельность, координированная работа сердечнососудистой и
дыхательной систем, отмечено также повышение давления (Василевская, 1971;
Замбржицкий, 1972; Беллер, 1977).
Согласно гипотезе П.В. Симонова (1981) в
медиобазальных отделах лобной коры происходит интеграция сигналов из внутренней
среды организма с двигательной активностью.
П.В. Симонов
(1981) рассматривая лобную кору как одну из структур, участвующих в организации
поведения, отмечает, что при повреждении лобных долей эмоциональная сфера
человека нарушается сильнее, чем при поражении любых других областей мозга. Так
как возникновение эмоционального напряжения сопровождается переходом к иным,
чем в спокойном состоянии, формам поведения, принципам оценки внешних
сигналов
и реагирования на них, то реакции на внешние раздражители зависят от внутреннего
состояния, которое определяется, в частности, балансом активности симпатической
и парасимпатической системы.
Для выявления возможных изменений в
эмоциональной сфере, происходящих у животных с сочетанным повреждением ствола
мозга и симметричных орбитофрональных образований, оценивали соотношение
"спокойных" и "эмоциональных" крыс до и после операции
(рис.7.5). Эти данные сопоставлялись с аналогичными данными у крыс с
изолированным стволовым повреждением.
Рис.7.5. Диаграмма, показывающая изменение
количества эмоциональных животных в группе с двусторонним повреждением
орбитофронтальной коры в сочетании со стволовым повреждением до операции
(внутренний круг) и после операции (наружный круг). 1 - спокойные животные; 2 -
эмоциональные.
Оказалось, что сочетанное стволовое и
двухстороннее орбитофронтальное
повреждение вызывает увеличение числа эмоциональных
животных.
Однако, по критерию хи-квадрат (%2=2.25; p
< 3.84) эти изменения, хотя и
превосходящие значимость однонаправленных сдвигов в группе с
изолированным
стволовым повреждением, могут рассматриваться лишь в
качестве
тенденции.
В данной группе не было выявлено специфических
изменений в эмоциональной сфере
по сравнению с изолированным стволовым повреждением. Эмоциональные реакции
носили пассивно-оборонительный характер.
Сравнивая изменения в эмоциональной сфере у крыс
с разными исходами, следует обратить внимание на отсутствие эмоциональных
реакций до операции у большинства крыс с неосложненным и осложненным течением послеоперационного
периода. После операции у всех этих крыс появились эмоциональные реакции. А у
всех животных с летальным исходом до операции наблюдались эмоциональные
реакции. После операции эмоциональные реакции в большинстве случаев сохранились
на дооперационном уровне, но у двух крыс (№70, №71) в послеоперационный период
эмоциональные реакции исчезли и отсутствовали до момента гибели животных.
Появление агрессивных реакций, как при изолированном повреждении ствола, перед
гибелью животных нами отмечено не было.
Следует обратить внимание на то, что в группах с
летальным исходом, как при изолированном повреждении ствола, так и с сочетанным
стволово-орбитофронтальным разрушением число крыс с исходно повышенной
эмоциональностью преобладало и в последнем случае достигло максимума.
Глава 8. Особенности латерализованного
повреждения орбитофронтальной коры на фоне острого стволового патологического
очага
Учитывая парную организацию ОФК, мы посчитали
целесообразным проанализировать особенности протекания послеоперационных
адаптивных реакций мозга у крыс со стволово-орбитофронтальным повреждением,
сочетанным с разной латерализацией повреждения ОФК.
Если функциональная асимметрия подкорковых
структур у человека и животных получает прямое подтверждение сравнительно
недавно (Брейже, 1967; Левшина с соавт., 1977; Glick
et al.,
1977; Лапина с соавт., 1984; Квирквелия, 1987; Ванециан, 1990, 1993; Chida,
Toyosawa, 1992; Чилингарян,
Богданов, 1998; Молодцова, 1999; Павлова, 1998; Sullivan,
Gratton, 1999, 2002; Concha,
Wilson, 2001; Verstynen
et al.,
2001; Чилингарян, 2002; Ванециан, Павлова, 2003; Maguire,
Frith, 2003; Артюхина,
Саркисова, 2004; Pedraza
et al.,
2004), то исследования асимметрии больших полушарий головного мозга имеют
давнюю историю.
Впервые более 140 лет назад нейрохирург Ж.П.
Брока продемонстрировал доминантность левого полушария по речевой функции
человека. Сейчас общепризнано, что латеральная специализация больших полушарий
является одной из основных психофизиологических характеристик. Если первоначально
господствовало представление о тотальном доминировании левого полушария в
формировании высших психических функций, то сейчас склонны говорить о частичном
доминировании каждого из полушарий (Бианки, 1985, 1989) или специфичности
правого и левого полушарий по отношению к разным функциям. Хомская
рассматривает эту специфичность как относительную доминантность левого
полушария у человека (Хомская, 1987). В основе нейрофизиологического
обеспечения ФМА лежат, по-видимому, выявленные у человека полушарные различия
биоэлектрических процессов (Жаворонкова, 1999), а также особенности
функционального взаимодействия правого и левого полушарий со структурами ствола
(Шарова, 1999) и лимбико-диэнцефального комплекса (Болдырева с соавт., 2000).
Интенсивные исследования последних десятилетий
показали наличие функциональной асимметрии больших полушарий и у животных.
Вопрос о том, является ли эта асимметрия видовой или индивидуальной
окончательно не решен (Адрианов, 1986). В последнее время вокализация у птиц,
предпочтение конечности и пространственно-зрительная функция у крыс
рассматриваются как одно из проявлений видовой асимметрии (Walker,
1980; Danenberg,
1981; Бианки, 1985; Вартанян, Клементьев, 1991). Остается неясным вопрос об
относительной или абсолютной доминантности правого и левого полушария животных,
а также асимметрии в пределах разных функциональных систем.
Изучение моторной асимметрии у разных видов
животных фактически и позволягт говорить о доминантности того или иного
полушария. Такая асимметрия выявлена, в частности, у крыс при выработке
одностороннего манипуляторного навыка. Считается, что асимметрия в значительной
степени организуется в процессе обучения - впрочем, на фоне исходного
предпочтения используемой конечности (Сташкевич, 2000, Иоффе с соавт., 2002). Роль
обучения (в частности, сенсорного) подчеркивается также в работе Удаловой и
Михеева (1982), исследовавших формирование у крыс предпочтения направления
движения.
В некоторых работах последних лет функциональная
асимметрия полушарий связывается с особенностями биохимии правого и левого
полушарий. Так, по данным Авалиани с соавт. (2000), односторонняя травма мозга
крыс-матерей вызывает сдвиг равновесия между право- и левосторонними пептидными
факторами в структурах нейроэндокринной системы потомства, что, возможно, и
приводит к функциональной моторной асимметрии.
Вартанян и Клементьев (1991) показали наличие
генетически детерминированных видов химической асимметрии полушарий мозга
животных. Авторы постулируют существование различных химических маркеров правой
и левой половины головного мозга, к числу которых они относят, например,
эндогенные олигопептиды. По их мнению, активность олигопептидов отчетливо
проявляется в условиях органической патологии ЦНС и играет существенную
(возможно даже ведущую) роль в компенсаторном процессе.
Анализируя имеющиеся в литературе данные, мы
пришли к выводу, что особенности протекания компенсаторных процессов при разной
латерализации повреждения мозга человека и животных исследованы недостаточно,
при всей важности этой проблематики в теоретическом и практическом плане.
Как показано в предыдущей главе, при сочетанном
повреждении ствола с симметричными областями орбитофронтальной коры значимо
возрастало количество животных с летальным исходом. Однако вклад левой и правой
части этого симметричного образования в процессе компенсации после стволового
этапа операции, оставался не выясненным.
Реальными предпосылками к такой постановке
вопроса являлись и факты, отмеченные в главе 7: наиболее драматический исход
повреждения мозга имел место в тех случаях, когда значительное разрушение
ствола на уровне ВЯД сочеталось с двухсторонним, негрубым правосторонним либо
обширным левосторонним повреждением ОФК.
Была проведена серия специальных исследований, в
которых у 21 крысы за 7 дней до электрокоагуляции ствола на уровне ВЯД
производилось одностороннее электролитическое повреждение ОФК справа (11
животных) или слева (10 животных). У одной крысы (№69) повреждение ОФК было
механическим, обусловленным вживлением электрода для регистрации ЭА.
Схема экспериментов с оценкой особенностей
состояния животных до и после операции была стандартной (см. главы 5-7).
Послеоперационная динамика состояния
В таблице 8.1. и на соответствующих круговых
диаграммах представлено распределение крыс с латерализованным поражением ОФК с
сочетанным повреждением ствола мозга по исходам экспериментальной патологии -
при разной степени выраженности у них неврологических отклонений и сроках их
редукции.
Таблица 8.1. Распределение крыс по исходам с
разной степенью выраженности неврологических отклонений и срокам их редукции
Повреждение
Исход
Орбитофронтальная
кора справа + ствол
Орбитофронтальная
кора слева + ствол
Кол-во
Сроки
редукции «винта»
Кол-во
Сроки
редукции «винта»
Неосложненный
1
1
Осложненные
5
4+1w
7
8
3+5w
7;
9; 9; 44; 7
Летальный
5
1
Всего
11
10
Примечание: w
-
крысы линии Wistar
Выявлено, что при левостороннем повреждении ОФК
из 10 животных лишь у одного (№2112) не наблюдалось неврологических
послеоперационных нарушений. У 6 крыс имело место осложненное течение
послеоперационного периода с наличием неврологических отклонений в виде
появления винтообразных движений, наклонов туловища и головы в оперированную
сторону. Окончательная редукция этих неврологических отклонений не произошла
вплоть до усыпления крыс эфиром на 122-е сутки после операции. У трех особей
повреждение мозга закончилось летальным исходом: одна крыса (№115) погибла в
первые сутки после стволового этапа операции, две другие на 40-е и 130-е сутки.
Следует отметить, что гибель крыс с левосторонним
повреждением ОФК в поздние сроки после повреждения ствола мозга не
сопровождалась потерей веса. Не наблюдались трофические нарушения (в форме
сукровичных подтеков вокруг глаз, некротических пятен около рта и загрязнением
шерсти), как это имело место у животных, погибших после двухстороннего
разрушения ОФК в сочетании со стволовым повреждением. По данным
патологоанатомического исследования, гибель этих крыс не была связана с
нарушением системного кровообращения и не была непосредственно обусловлена
оперативным вмешательством. Поэтому при анализе данных указанные особи (крысы
№113, №124) были отнесены в группу с осложненным течением послеоперационного
периода.
При правостороннем повреждении ОФК у 5 из 11
животных имело место осложненное течение послеоперационного периода с
неврологическими нарушениями разной степени тяжести. Одна крыса была отнесена к
группе «неосложненных». Пять животных погибли, причем двое - в первые сутки
после операции.
В патологоанатомической картине гибели этих
животных (наступившей в период от первых до 7-8-х суток после стволового этапа
операции) выявлены некоторые особенности. У крыс № 114 и №109 ОФК была
повреждена справа. Смерть животных наступила вследствие гемотампонады
желудочков. Крыса №115 с левосторонним повреждением ОФК погибла в первые сутки
после экспериментального повреждения мозга. Было обнаружено, что гибель этого
животного
наступила, скорее всего, вследствие нарушения оттока крови от правого полушария
из-за закупорки вен справа.
У выживших крыс с правосторонним повреждением
ОФК при осложненном послеоперационном течении обращало на себя внимание
отсутствие винтообразных движений у выживших животных. Наличие «винта» было
прогностически неблагоприятным признаком при правостороннем повреждении ОФК
(табл.8.1.).
Из таблицы 8.1., в частности, следует, что в
группе с левосторонним повреждением ОФК все пять животных с винтообразными
движениями выжили. В то же время в группе с правосторонним повреждением ОФК из
трех крыс с винтообразными движениями погибли две (№62, №69). Статистический
анализ показал, что доля животных с наличием выраженного «винта» в первые дни
после операции доля крыс, выживших после операции на левом полушарии,
статистически достоверно отличается от доли крыс, выживших после операции на
правом полушарии (по точному критерию Фишера р = 0.048).
Рис. 8.1 Диаграмма распределения животных по
исходам с повреждением ствола в сочетании с право- (А) и левосторонним (Б)
повреждением орбитофронтальной коры в процентах. 1-
осложненное течение; 2 - летальный исход; 3 - легкое течение послеоперационного
периода.
На диаграмме 8.1. представлено распределение
животных по исходам при разрушении ствола в сочетании с правосторонним (А) и
левосторонним (Б) орбитофронтальным повреждением. Видно, что число особей с
летальным исходом преобладает в группе с правосторонним повреждением ОФК. В то
же время число животных с осложненным течением послеоперационного периода
больше в группе с левосторонним повреждением орбитофронтальной коры.
Проверка существенности различия числа умерших
животных при операции на левой и правой орбитофронтальной коре (без учета
степени выраженности послеоперационных неврологических нарушений) показало лишь
тенденцию к преобладанию доли погибших крыс при повреждении правого полушария
по сравнению с левым (по точному критерию Фишера р = 0.09). В данном случае
отсутствие достоверной разницы может быть обусловлено малым количеством
наблюдений.
Рассматриваемые формальные показатели отражают
далеко не полную картину послеоперационных событий. Так как
патологоанатомическими исследованиями было установлено, в частности, что и при
левосторонней деструкции ОФК гибель животных в ранние сроки после операции
бывает обусловленной изменениями, происходящими в правом (т.е. интактном) полушарии
(крыса №115). Таким образом, данные патологоанатомического исследования
являются дополнительным подтверждением значимости правой ОФК в выживании
животных после стволового повреждения.
Вместе с тем, сравнение сроков редукции
неврологической симптоматики у оперированных животных не выявило достоверных
различий между группами с право - и левополушарным повреждением ОФК.
Из данных литературы известно, что при патологии
головного мозга человека (тяжелая черепно-мозговая травма, сосудистые
поражения) длительность и успешность восстановления зависят как от стороны
поражения мозга, так и от индивидуального профиля их функциональной
межполушарной асимметрии (Жирмунская, 1989; Доброхотова, Брагина, 1993). Это
обусловлено, в частности, тем, что на разных стадиях восстановления
церебральных функций имеет место попеременное преимущественное включение в
компенсаторный процесс правого или левого полушария (Болдырева с соавт., 1993).
Причем, доминирование активности одного из полушарий бывает обусловлено, в том
числе и функциональным состоянием лимбико-диэнцефальных и стволовых
регуляторных структур мозга (Брагина, Доброхотова, 1988; Шарова с соавт., 1993,
1995; Болдырева с
соавт., 2000).
Известно, что для человека справедливо наличие
т.н. видовой асимметрии: для большинство людей характерна праворукость. Вопрос
наличие видовой асимметрии у крыс остается дискуссионным. Все экспериментальные
животные, с которыми мы работали, делились на три приблизительно равные группы:
правшей, левшей и амбидекстров, Кроме того, встречались животные,
"рукость" которых определить не удавалось. В исследованных нами
примерах экспериментальной патологии сохранность правой ОФК оказалась более
существенной для выживания при стволовых левосторонних повреждениях по всей
группе оперированных животных. Правомерен вопрос о том, зависит ли протекание
восстановительных процессов у крыс от наличия и типа функциональной
межполушарной асимметрии и от ее характера.
Напомним, что показателями моторной
функциональной асимметрии служили «рукость», а также направления движения в
открытом поле. Причем предпочтение по одному тесту не всегда совпадало с
таковым по другому. По данным литературы, эти функции могут иметь разные
системы обеспечения. В основе пространственного предпочтения лежат
индивидуальные различия в концентрации дофамина в правой и левой части
нигростриарной системы, значимость соотношения активности между ними, а также
особенности корково-нигростриарного взаимодействия (Glick,
Cox, 1976; Glick
et al,
1977; Danenberg,
1981; Бианки с соавт. 1982; Бианки, Филиппова, 1984).
В нашей работе мы попытались сравнить характер
протекания восстановительных процессов с типом функциональной межполушарной
асимметрии. В таблице 8.2. приводится распределение крыс с разными исходами
сочетанного стволового и летерализованного орбитофронтального повреждения по
моторной функциональной асимметрии и предпочтению направления
движения в открытом поле.
Таблица 8.2 Распределение крыс с разными
исходами по моторной функциональной асимметрии и предпочтению направления движения
Сторона
Повреждение
справа
Повреждение
слева
Исход
лапа
Неосложн
Осложненный
Летальный
Неосложн.
Осложненный
летальный
I
II
III
I
II
III
Правая
3
1
Левая
1
5
Амбидекстр
2
1
3
1
Не определена
1
1
3
Направо
3
1
2
1
Налево
1
1
3
2
3
Нет предпочтения
2
2
3
Оказалось, что для животных, которые в открытом
поле предпочитали движение налево, то есть с правосторонним доминированием по
этому показателю, критичным было сохранение целостности правой ОФК. Для
животных с левосторонним доминированием важна была целостность левой ОФК. Таким
образом, исходя из наших данных, для выживания крыс со
стволово-орбитофронтальным повреждение преимущественное значение имела
сохранность полушария, контралатерального (возможно доминирующего) по функции
"направление движения в открытом поле". Но в связи с недостаточностью
выборки данное утверждение нуждается в дальнейшей проверке.
По показателю "рукости" не было
выявлено четких различий между право- и левополушарными животными. Интересным
представляется тот факт, что из 6 погибших в обеих группах животных две крысы
(№114, №115) были амбидекстрами, а у трех не удалось определить сторону
предпочтения. И только одна крыса (№69) была левополушарной (табл. 8.2). Эти
данные можно рассматривать с точки зрения стрессоустойчивости организма, так
как индивидуальные психологические качества личности определяют устойчивость
или склонность к психосоматическим заболеваниям (Юринов, 2001). А у крыс
межполушарная асимметрия выражена в большей степени у животных с сильным типом
нервной системы. Крысы со слабым типом нервной системы, менее
стрессоустойчивые, не имеют видоспецифической межполушарной асимметрии (Мурик,
1990).
Изменения в эмоциональной сфере
Учитывая значимость передних отделов новой коры
в процессах мотивациий и эмоций (Наута, 1963; Симонов, 1981), а также данные
литературы о неравнозначности правого и левого полушарий в отношении
формирования положительных и отрицательных эмоций (Лурия, Симерницкая, 1977;
Симонов, 1981, 1999; Blonder
et al.,
1991; Adolphs
et al.,
1996; Михайлова с соавт., 1998; Преображенская, 2000; Павлова, 2001; Harmon-Jones,
2003), представляли интерес и изменения в эмоциональной сфере после повреждения
ствола в сочетании с повреждениями ОФК правого или левого полушария.
Сопоставление эмоциональности крыс до и после
операции проводили по описанной уже методике (см. главы 5-7). Полученные данные
свидетельствуют об определенных различиях в степени выраженности эмоциональных
реакций в зависимости от латерализации повреждения ОФК (рис. 8.2. - круговые
диаграммы). При повреждении ствола в сочетании с повреждением правой ОФК
количество эмоциональных животных увеличилось на 16% (с 67% до 83%). Эти
изменения были недостоверны по критерию
хи-квадрат. Однако, в случае с левосторонним повреждением ОФК эмоциональность
среди всех оперированных животных возросла на 63%, что
было
статистически значимым ( %2=5; р> 3.84)
Рис.8.2 Диаграмма, показывающая изменение
количества эмоциональнореактивных
животных до операции и после сочетанного повреждения ствола с левосторонним (А)
и правосторонним (Б) повреждением орбитофронтальной коры.
1
- спокойные животные; 2 - эмоционально реактивные животные
При сочетанном повреждении ствола с ОФК справа и
слева преобладали реакции пассивно-оборонительного типа, за исключением крысы
№127 с левосторонним повреждением ОФК. Эта крыса была отнесена в группу с
осложненным
течением послеоперационного периода, хотя у нее наблюдался после незначительный
(до 30°) наклон головы. Однако этот дефект наблюдался до 119-ти суток, вплоть
до усыпления животного. У крысы №127 мы наблюдали агрессивно-оборонительные
реакции в течение тридцати суток операции на стволе, которые затем также
сменились пассивно- оборонительными реакциями
Таким образом, в настоящем разделе работы
показано, что стволово-орбитофронтальное повреждение приводит к значительному
нарастанию тяжести состояния
животных и высокой вероятности летального исхода в первые две недели после
операции.
По данные патоморфологического обследования, гибель
животных при стволово-орбитофронтальном повреждении была обусловлена нарушением
мозгового кровообращения по геморрагическому типу и, вероятно, вторичной
дисфункцией гипоталамуса. Гибель животных наступала чаще при двухсторонней либо
правосторонней деструкции ОФК. Причем, в большинстве случаев у погибших крыс
выявлялось правополушарное доминирование по направлению движения в открытом
поле: из пяти погибших животных три крысы предпочитали движение влево. Еще у
двух не было выявлено какого-либо предпочтения направления движения. Но в связи
с недостаточностью выборки мы не можем говорить о достоверности полученных
результатов.
Объяснить полученные данные о связи повреждения
ОФК и исходом возможно, по-видимому, опираясь на информацию об активной роли
передних отделов полушарий в висцеральной регуляции. По данным ряда авторов,
двусторонняя цингулоэктомия у животных вызывает нарушения деятельности
внутренних органов. Происходит, в частности, гиперактивация блуждающего нерва и
парасимпатических веточек глазодвигательного и лицевого нерва. При этом
усиливается моторика желудочно-кишечного тракта, наблюдается расширение
зрачков, гиперсекреция слюны и так далее. Эти проявления связываются с
нарушением синергии парасимпатической и симпатической систем (Smith,
1945; Ward, 1948; Showers,
1959; Lofwing,
1961; Alanis
et al.,
1966; Замбржицкий, 1972; Вейн, Соловьева, 1973; Беллер, 1977; Карцева, 1985; Burns
et al.,
1985; Баклаваджян с соавт., 1987, 2000; Багаев, Пантелеев, 1995).
Ряд авторов (Замбржицкий,1972; Nauta,
1972; Беллер, 1977) рассматривает лобную область как неокортикальное
продолжение лимбической системы, при разрушении и стимуляции которой у разных
видов животных наиболее часто наблюдаются угнетение некоторых вегетативных
функций. Отмечено нарушение сердечной деятельности, координированной работы
сердечно-сосудистой и дыхательной систем, а также повышение давления (Kaada
et al.,
1949; Василевская, 1971; Замбржицкий, 1972; Беллер, 1977).
Сравнение экспериментальных данных с данными,
полученными в условиях клиники, подтверждает универсальность функций переднего
отдела поясной извилины (поле 24). Так, у больных с опухолью, расположенной в
большой щели мозга, имелись симптомы нарушения вегетативной нервной регуляции и
поведения. Имела место повышенная агрессивность и плаксивость (Замбржицкий,
1972; Ongur, Price,
2000).
По мнению Симонова (1981), в медиобазальных
отделах лобной коры происходит интеграция сигналов из внутренней среды
организма с двигательной активностью. Поэтому повреждение передних отделов
приводит к нарушению зоосоциального поведения крыс. Кроме того, повреждение
лобных долей у человека приводит к наибольшим (по сравнению с другими областями
мозга) нарушениям в эмоциональной сфере. Наши исследования показали, что
повреждение лобных отделов полушарий у крыс также сопровождается выраженными
эмоциональными нарушениями, которые значимо больше проявлялись при
левостороннем повреждении ОФК.
Заключение
В настоящей работе нам удалось в эксперименте
получить долго живущую "модель" острого поражения ствола посредством
его электролитической коагуляции на уровне латерального вестибулярного ядра
Дейтерса. Сопоставление клинико-поведенческих последствий такого повреждения в
наших экспериментах с данными литературы (Сепп соавт., 1950; Чернышев, 1983;
Благовещенская, 1990) свидетельствует об определенном сходстве полученных
двигательных и вестибулярных расстройств у крыс с проявлениями стволовой
патологии у человека, что обусловлено определенной универсальностью организации
этих функций у млекопитающих на уровне ствола. Наблюдение за состоянием
животных после электролитической стволовой коагуляции и распределение их по
трем группам в зависимости от особенностей динамики состояния и исходов также
указывает на определенные аналогии с вариантами послеоперационного течения у
нейрохирургических больных после удаления стволовой опухоли (Баркалая, 1991;
Шарова, 1991, 1993; Брагина с соавт. 1997).
Все эти обстоятельства свидетельствуют о
правомочности использования предложенной модели для изучения
адаптивно-компенсаторных перестроек ЦНС при острой стволовой патологии.
Важным представляется тот факт, что
послеоперационное течение и исход стволового повреждения у животных не зависели
только от объема стволового разрушения, а определялось многими факторами, к
числу которых относятся реакции ткани и сосудов мозга на электротравму,
целостные церебральные реакции на повреждение и др. Выявлена, в частности,
значимость исходной эмоциональности животных, а также предпочитаемого
направления движения в открытом поле (раздел I).
Данные динамического анализа ЭА у животных с
локальным стволовым повреждением также демонстрируют значительное сходство с
изменениями ЭЭГ у больных с опухолевым стволовым повреждением в раннем
послеоперационном периоде. И у животных, и в клинических наблюдениях послеоперационная
нейродинамика носит фазный характер с последовательным повышением сочетанности
определенных зон мозга; частота доминирующей синхронизации биопотенциалов
тета-диапазона (6-7 или 4-5 Гц) различается при благоприятном и неблагоприятном
вариантах послеоперационного течения (Гриндель с соавт., 1983; Шарова с соавт.,
1991, 1993). Совпадение временных и частотных параметров биоэлектрических
процессов, наблюдаемых в условиях локального стволового повреждения у животных
и человека, служит еще одним подтверждением физиологической общности
(универсальности) церебральных механизмов гомеостатической (висцеральной)
компенсации мозга млекопитающих. Тем важнее представляются результаты
экспериментального исследования полушарно-гиппокампальных и стволово-гиппокампальных
отношений у животных, проведение которого невозможно у человека.
Полученные экспериментальные данные позволяют
ответить на ряд вопросов, возникающих при трактовке послеоперационных
ЭЭГ-изменений у человека.
. Тождество поведения тета-составляющих ЭА мозга
крыс и ЭЭГ человека при остром стволовом повреждении служит объективным
доказательством генетического единства этого вида активности и подтверждает
важную роль гиппокампа в его происхождение (Arnolds
et al.,
1979; Bland, 1986; Haas,
1987]. 2.При отсутствии единого представления о механизмах генерации тета-ритма
можно высказать нескольких предположений о нейрофизиологических механизмах
формирования разночастотной тета-активности, которая прогностически значима для
острых патологических состояний человека: частота тета-ритма может отражать как
разное функциональное состояние одной и той же функциональной системы, так и
разную организацию этой системы при разных типах стволового патологического
очага (стойкий или углубляющийся). Наши экспериментальные электрофизиологические
исследования показали, что частотные параметры синхронизации тета-активности
(4-5 или 6-7 Гц) определяются, прежде всего, состоянием
стволово-гиппокампального входа. Частота снижалась по мере угнетения
функциональной активности ствола (Бражник, Виноградова, 1983). Кроме того,
возможно перераспределение активности в пределах стволовой медиаторной системы
(Меликов, 1987, Vertes
et al.,
1993). Полученные данные подтверждают скорее предположение о разном состоянии
одной и той же функциональной системы.
Когерентный анализ ЭА позволил как бы
«визуализировать» особенности динамики стволово-гиппокампального и
гиппокампально-кортикального взаимодействия при различных состояниях
ретикуло-септального входа. Показано, например, что в условиях относительной
морфологической сохранности ствола именно этот вход является «запускающим
звеном» синхронизации на частоте 6-7 герц. Если же состояние активирующей
стволовой системы резко изменено (угнетено) вследствие значительного объема
стволового повреждения на уровне ВЯД (осложненное течение), лидирующую роль в
запуске этой реакции могут на некоторое время брать передние отделы коры и
гиппокамп. При летальном исходе показана возможность формирования автономной
синхронизированной гиппокампально-корковой системы на частоте 4-5 Гц, которая,
вероятно, сопряжена с относительной стабилизацией состояния низкоуровневого
висцерального обеспечения.
Данные наших электрофизиологических исследований
подтверждают участие ОФК в формировании системной приспособительной реакции при
остром стволовом повреждении. Ранняя послеоперационная стволово-гиппокампальная
синхронизация биопотенциалов на частоте 6-7 Гц с последующим вовлечением
передних отделов коры характерна для выживших крыс. У погибших животных падение
межполушарных связей ОФК предшествовало летальному исходу. Следует отметить,
однако, что у человека роль передних отделов полушарий в послеоперационных
адаптивных перестройках более существенна (Шарова с соавт., 1993), что связано,
по-видимому, с прогрессивным морфо-функциональным развитием лобных отделов
мозга в филогенезе.
При сравнении последствий сочетанного поражения
мозга в наблюдениях с разной его топографией обращает на себя внимание
существенные различия в исходах между группами со стволово-гиппокампальными и
стволово-орбитофронтальными повреждениями.
Стволово-гиппокампальная коагуляция усугубляет
по большей части стволовую неврологическую симптоматику и пролонгирует время ее
обратного развития, т. е. вызывает подкрепление очага стойкой патологической
активности. Этот факт соответствует данным литературы (Русинова, 1989) о
подкреплении доминантных очагов, сформированных в других отделах ЦНС, путем
раздражения гиппокампа. Стволово-орбитофронтальное повреждение приводит к
значительному нарастанию степени тяжести состояния животных и высоковероятному
летальному исходу в первые две недели после операции.
К числу послеоперационных различий можно отнести
также ряд специфических особенностей состояния животных. При стволово-ОФК
повреждениях наблюдалось появление сукровичных подтеков вокруг глаз и резкое
падение веса в первые дни после операции, которое было особенно выражено у
животных с летальным исходом. Снижение веса в группе с летальным исходом
достоверно отличалось от падения веса в других группах животных (см.график на стр.
137). Потеря веса при стволово-гиппокампальном повреждении менее значительна.
Данные патоморфологического обследования
погибших животных указывают на разные причины летального исхода в случае разных
вариантов сочетанного повреждения мозга при равнозначности общего объема
коагуляции. Гибель крыс при стволово-гиппокампальном разрушении, как правило,
была вызвана общим снижением уровня иммунной защиты организма, так как
экспериментальное воздействие на структуры дорсального и вентрального
гиппокампа приводит к изменению иммунного ответа организма (Магаева, Морозов,
2005). Стволово-орбитофронтальное повреждение вызвало гибель животных при
значительном разрушении латерального вестибулярного ядра Дейтерса, либо
кохлеарного ядра с затеканием тока на ядро Дейтерса в сочетании чаще с
двусторонним, а также негрубым правосторонним либо обширным левосторонним
повреждением орбитофронтальной коры. И в данном случае гибель была обусловлена
нарушением мозгового кровообращения по геморрагическому типу и, вероятно, вторичной
дисфункцией гипоталамуса. При этом повреждения в стволе наряду с двусторонним
или с односторонним разрушением ОФК, но без вовлечения ядра Дейтерса не
приводила к столь гибельным для животных последствиям. Тяжесть последствий
можно объяснить участием именно этого отдела коры в висцеральной регуляции.
Сочетанное повреждение ствола и ОФК вызывает столь мощные нарушения
гемодинамики мозга, что это, в конечном счете, увеличивает вероятность гибели
животных.
Гибель животных с изолированным стволовым
повреждением не была обусловлена нарушениями мозгового кровообращения, не
сопровождалась потерей веса, не отмечалось также дисфункций иммунной системы,
как в случае сочетанных повреждений.
Клинические наблюдения показали неодинаковые
функциональные возможности правого и левого полушарий (Мосидзе с соавт., 1977;
Жирмунская, 1986; Брагина, Доброхотова, 1988; Wittling
W., 1990; Хомская с
соавт., 1997). В настоящее время взаимодействие полушарий головного мозга
рассматривается как взаимодополняющее, взаимокомпенсирующее в реализации
различных функций центральной нервной системы. Такая морфофункциональная
организация мозга оказывается целесообразной для развития компенсаторного
процесса при возникновении латерализованного патологического процесса (Советов,
1988; Кураев, Сороколетова, 1996). Выяснение вклада ОФК правого и левого
полушария и правого и левого гиппокампа показали, что значение правого и левого
полушарий в компенсаторных процессах четко видно именно на уровне ОФК, но не
гиппокампа.
При стволово-орбитофронтальном варианте
летальный исход наиболее вероятен при двустороннем или правостороннем
повреждении ОФК. С одной стороны эти данные подтверждают выявленные в клинике
факты о большей причастности правого полушария к процессам висцеральной
регуляции
(Доброхотова, Брагина, 1977; Жаворонкова, 1990;
Болдырева с соавт., 1993; Жаворонкова, Добронравова, 1993; Шарова с соавт,
1993). Наряду с этим, можно предположить большую значимость правого полушария
как контралатерального поражению ствола на уровне ВЯД слева (Советов, 1988).
При стволово-гиппокампальном повреждении связь исхода с латерализацией
разрушения гиппокампа оказалась не очевидной.
Следует подчеркнуть, что моделирование
сочетанного стволово-гиппокампального и стволово-орбитофронтального повреждения
у животных может иметь самостоятельное значение в качестве модели
травматического повреждения мозга, т.к. 1) при черепно-мозговой травме
повреждения мозга всегда множественны; 2) состояние ствола играет определяющую
роль в ее исходе; 3) наиболее вероятны глубокие разрушения лобной и височной
локализации (в том числе и латерализованные).
Наконец, анализируя стволово-лимбические
взаимоотношения, нельзя не коснуться вопроса о связи их с эмоциональными
проявлениями. В нашей работе мы затрагивали его в двух аспектах: 1) влияет ли
дооперационная эмоциональность животных на исход локального стволового
поражения; 2) имеется ли специфический эффект повреждения исследуемых
лимбических образований (гиппокампа и орбитофронтальной коры) в эмоциональной
сфере
Причем, второй аспект наблюдения вполне
правомерен, т.к. вопрос о степени участия отдельных лимбических структур в
формировании эмоциональных реакций разной интенсивности и знака до сих пор
окончательно не выяснен.
Учитывая тяжесть повреждения головного мозга, мы
могли рассматривать каждый из указанных аспектов лишь в ограниченной степени.
Тем не менее, были получены вполне определенные результаты.
. Выявлена статистически значимая зависимость
исхода стволового повреждения от дооперационной эмоциональности животных: число
крыс с исходно повышенной эмоциональностью проградиентно нарастает от
неосложненного
к осложненному течению и летальному исходу - при наличии достоверной, хотя и
нелинейной, связи между этими показателями (р = 0.08). Отмечается появление
эмоциональных реакций пассивно оборонительного типа перед улучшением
неврологических показателей в случае благоприятного исхода у животных с
осложненным течением послеоперационного периода, а также агрессивных реакций
перед гибелью животных.
Рис. 9.1 Увеличение количества эмоциональных
животных в группах с разными типами повреждений. По оси абсцисс номера групп:
1-изолированный ствол; 2 - ствол и билатеральное повреждение гиппокампа; 3
-ствол и билатеральное повреждение ОФК; 4 - ствол и ОФК справа; 5, 6 - ствол и
гиппокамп слева и справа соответственно; 7 - ствол и ОФК слева. По оси ординат
- проценты. * р > 3.84
2.Показано наличие специфических эффектов в
эмоциональной сфере при повреждении орбитофронтальной коры или гиппокампа в
сочетании с повреждением ствола мозга. В рамках настоящего исследования были
проанализированы "эмоциональные эффекты" семи вариантов органического
поражения головного мозга: изолированное стволовое (на уровне левого
вестибулярного ядра Дейтерса), сочетанное повреждение ствола и двухстороннего
гиппокампа, ствола с право- и левосторонним повреждением гиппокампа, ствола с
двухсторонним повреждением ОФК, а также ствола с право- и левосторонним
повреждением ОФК. Углубленный статистический (в том числе и по методу LSD
(least significant
difference) анализ
соотношения "спокойных" и "эмоциональных" животных до и
после операции в этих 7 группах показал, что можно выделить два варианта "
эмоциональных последствий" повреждения головного мозга (рис. 9.1). При
первом варианте (изолированное повреждение ствола, сочетанное повреждение
ствола с билатеральным повреждением гиппокампа, орбитофронтальной коры или
повреждением орбитофронтальной коры справа) наблюдается незначимое увеличение
доли "эмоциональных" животных после операции по сравнению с
дооперационным уровнем. При втором варианте (повреждение ствола в сочетании с
правым или левым гиппокампом, или орбитофронтальной коры слева) наблюдается
значимое увеличение доли "эмоциональных" животных после операции,
наиболее резкое для случаев сочетанного поражения ствола с левой ОФК, либо с
правым гиппокампом. Таким образом, можно думать, что в рамках общей
неспецифической тенденции к послеоперационному усилению эмоциональности
животных, максимальные сдвиги в эмоциональной сфере связаны с системами
"ствол - левая ОФК" и "ствол - правый гиппокамп". Более
четко трактовать выявленные тенденции сложно: это могут быть как эффекты
раздражения повреждаемых лимбических структур, так и снятие тормозных влияний
вследствие их выключения.
Нам кажется важным обратить внимание на исход
при латерализованном повреждении ОФК и максимальных сдвигах в эмоциональной
сфере при повреждении слева. Возможно функции мобилизации энергетических
ресурсов (Нуцубидзе, 1969; Хананашвили, 1972, 1983; Симонов, 1993, 2001),
которая, как полагают, свойственна эмоциям, играет немаловажную роль в
компенсаторно-восстановительных процессах.
Выводы
1. Анализ
последствий изолированного электролитического разрушения ствола мозга крыс на
уровне латерального вестибулярного ядра Дейтерса мозга крыс вызывающего
воспроизводимый и стойкий характер двигательных и вестибулярных неврологических
нарушений, свидетельствует о правомерности использования предложенной
экспериментальной модели для изучения компенсаторных механизмов при стволовой
патологии человека.
2. Исследование
электрической активности мозга крыс (с использованием спектрально-когерентного
анализа) до и после изолированного стволового повреждения
позволило
"визуализировать"
послеоперационную динамику стволово-гиппокампального и гиппокампально-полушарного
взаимодействия и выявить зависимость этой динамики от состояния
ретикуло-септального входа.
3. Нейрофизиологические
особенности протекания системных приспособительных реакций мозга крыс,
развивающихся при одностороннем локальном стволовом повреждении, наиболее ярко
отражаются в изменениях пространственно-временной организации тета-активности
орбитофронтальной коры, гиппокампа и интактного ствола: а) ранняя
стволово-гиппокампальная синхронизация биопотенциалов на частоте 6-7Гц с последующим
вовлечением в этот процесс передних отделов коры характерна для выживших
животных; б) формирование автономной гиппокампально-полушарной системы
возбуждения на частоте 4-5 Гц предшествует летальному исходу.
4. Установлена
статистически значимая зависимость исхода локального повреждения ствола
головного мозга от уровня эмоциональности животных в дооперационном периоде:
крысы с исходно повышенной эмоциональностью более склонны к осложненному
развитию патологического процесса и летальному исходу.
5. Морфо-функциональные сопоставления
животных с идентичным по объему электролитическим стволовым,
стволово-гиппокампальным и стволово-орбитофронтальным разрушением позволили
выявить важную и специфическую роль гиппокампа и ОФК (составляющих лимбической
системы) в адаптивно-компенсаторных процессах ЦНС при повреждении ствола мозга:
а) двусторонняя или односторонняя
деструкция гиппокампа наряду со стволовым повреждением усугубляет по большей
части стволовую неврологическую симптоматику и пролонгирует время ее обратного
развития, т. е. вызывает подкрепление очага стойкой патологической активности
стволовой локализации;
б) двухсторонняя или преимущественно
правосторонняя деструкция ОФК наряду со стволовым повреждением приводит к
нарастанию тяжести состояния животных и более вероятному летальному исходу в
первые две недели после операции, что обусловлено расстройством мозгового
кровообращения по геморрагическому типу и, по-видимому, вторичной дисфункцией
гипоталамуса.
6. При общей неспецифической тенденции к
послеоперационному усилению эмоциональности животных наибольшие сдвиги в
эмоциональной сфере отмечаются при комбинациях повреждения "ствол - левая
ОФК" и "ствол - правый гиппокамп".
Литература
1.
Авалиани
Т.В., Огурцов Р.П., Пузырева В.П., Серякова О.Р. Латерализация травмы мозга у
крыс-самок (Вистар) определяет иммунный и неврологический статус потомства //
Росс.физиол.журн. 2000. Т.86.№12. - С.1565 - 1572.
2.
Адрианов
О.С., Боголепов Н.Н. Проблемы компенсаторно-восстановительных процессов при
органических и функциональных поражениях мозга. В кн.:
Функционально-структурные основы системной деятельности и механизмы
пластичности мозга. Сб.научн.тр., вып.ГУ, М. 1975.
3.
Адрианов
О.С. Проблема структурной организации правого и левого полушарий мозга.
В
сб.:
Нейропсихол.анализ
межполушарн.асим.мозга. Под ред. Е.Д.
Хомской.
М.:Наука, 1986. - С.9-13.
4.
Адрианов
О.С. О структурной организации интегративных функций мозга // Вестн.АМН, 1987,
№8. - С.28-35.
5.
Адрианов
О.С. Принципы организации церебральных функций и узловые механизмы поведения. В
кн.: О принципах структурно-функциональной организации мозга. Избранные научные
труды. М. 1999. - С.143-239.
6.
Айрапетянц
М.Г.
Экспериментальные
неврозы // Успехи физиол.наук. 1985. Т.16.
№2. - С. 8-32.
7. Айрапетянц М.Г. Участие церебральной
гипоксии в патогенезе неврозов (новая концепция) // Журн.высш.нерв.деят. 1997.
Т.47. №2.-С.412-419.
8.
Айрапетянц
М.Г., Вейн А.М. Неврозы в эксперименте и клинике. М.: Наука, 1982. - 271с.
9.
Айрапетянц
М.Г., Мехедова А.Я., Козловская М.М., Незнамов Г.Г. Моделирование невроза у
собак и изучение действия антидепрессантов // Журн.
высш.
нерв.
деят.
1986. Т.36. №6. - С.1131-1138.
10.Айрапетянц
М.Г., Хоничева Н.М., Мехедова А.Я., Х.Ильяна Вильяр. Реакции на умеренные
функциональные нагрузки у крыс с индивидуальными особенностями
поведения//Журн.высш.нерв.деят. 1980. Т.30. №5. - С.994-1002.
11.Анохин
П.К. Проблема компенсации нарушенных функций и ее значение для клинической
медицины // Хирургия. 1954, №10. - С.3-11.
12. Анохин П.К. Общие принципы компенсации
нарушенных функций и их физиологическое обоснование. В кн.: Труды научной
сессии по дефектологии. М.: Медицина. 1958. - С.54-55.
13. Анохин П.К. Проблема центра и
периферии в современной физиологии нервной деятельности. В кн.: Очерки по
физиологии функциональных систем. П.К. Анохин.
М.: Медицина, 1975, 448 с. (стр.63-107).
. Артюхина
Н.И., Саркисова К.Ю. Асимметрия структурных изменений в гиппокампе у крыс при
двусторонней окклюзии общих сонных артерий. Тез.конф. «Современное состояние
асимметрии мозга», 2000. - С.20-21.
. Артюхина
Н.И., Саркисова К.Ю. Межполушарная асимметрия повреждений гиппокампа после
двусторонней перевязки общих сонных артерий // Росс.физиол.журн. 2004. Т.90.№2.
- С.146 - 156.
. Аруин
Л.И. Органы пищеварения. В сб.: Структурные основы адаптации и компенсации
нарушенных функций. Ред. Д.С. Саркисов.
М.:
Медицина,
1987. - 448с.
. Асратян
Э.А. Лекции по некоторым вопросам нейрофизиологии. М.,изд.
АН
СССР, 1959.
. Бабаева
А.Г. Кроветворные и лимфоидные органы. В сб.: Структурные основы адаптации и
компенсации нарушенных функций. Ред. Д.С. Саркисов.
М.:Медицина, 1987. - 448с
. Бабияк
В.И., Ланцов А.А., Базаров В.Г.. Клиническая вестибулология: Руководство для
врачей.- СПб.: Гиппократ, 1996.- 336 с., ил.
. Багаев
В.А., Пантелеев С.С. Эффекты стимуляции лимбической коры на ответы нейронов
ядер ваго-солитарного комплекса, вызванные раздражением блуждающего нерва //
Докл. РАН, 1995. Т.340, № 4. -С.555-558
. Баклаваджян
О.Г., Еганова В.С., Каспарова С.Р. и др. Сравнительное исследование механизмов
изменения вазомоторной симпатической активности при раздражении различных
структур лимбической коры и гипоталамуса // Физиол.журн.СССР, 1987. Т.73. №3. -
С. 373-383.
. Баклаваджян
О.Г., Нерсесян Л.Б., Аветисян Э.А., Аветисян И.Н., Аршакян А.В., Багдасарян
К.Г., Еганова В.С., Погосян Н.Л. Нейронная организация лимбико-(цингуло-)
висцеральной рефлекторной дуги. Успехи физиол.наук. Т.31, №4, 2000.- С.11-23.
23. Баркалая
Д.Б. Клиника раннего послеоперационного периода у больных с опухолями задней
черепной ямки. Дисс. канд.мед.наук.
М.,
1991.
24. Батуев
А.С. Нейрофизиология коры головного мозга. Модульный
принцип
организации: Курс лекций. - Л.: ЛГУ, 1984. - 184 с.
25. Беленков
Н.Ю. Принцип целостности в деятельности мозга. М.:
Медицина,
1980, 309 с.
. Беллер
Н.Н. Висцеральное поле лимбической коры. - Л.: Наука, 1977. - 158 с..
. Березовский В.К. Стволовые пути
инициации локомоции. Нейрофизиология.
1991, Т.23, №4. - С.488-505.
28. Бехтерева
Н.П. Здоровый и больной мозг человека. Л.: Наука, 1988. -262с.
. Бианки В.Л. Индивидуальная и
видовая межполушарная асимметрия у
животных // Журн.высш.нерв.деят. 1979. Т.29. №2. - С.295-303.
30. Бианки
В.Л. Обзор: Латеральная специализация мозга животных //
Физиол.журн.
СССР, 1980. Т.66. №11. - С.1593-1607.
31. Бианки
В.Л. Асимметрия мозга животных. Л.: Наука, 1985, 295 с.
32. Бианки
В.Л. Механизмы парного мозга. - Л.: Наука, 1989- 264 с.
33. Бианки В.Л., Мурик С.Э.,
Филиппова Е.Б. Межполушарная асимметрия
положительных эмоциональных реакций у крыс // Журн.высш.нерв.деят.
1988. Т.38. №4. - С. 693-700.
34. Бианки
В.Л., Филиппова Е.Б., Пошивалов В.П. Межполушарная
асимметрия
двигательной и исследовательской активности у мышей //
Журн.высш.нерв.деят.,
1982. Т.32, 33. - С442.
35. Бианки В.Л., Филиппова Е.Б.
Межполушарная асимметрия и двигательная
активность у грызунов // Журн.высш.нерв.деят.. 1984.
Т.34.№6.
- С.1093-1099.
. Благовещенская
Н.С. Изменения электрической активности коры при опухолях задней черепной ямки
// Проблемы современной нейрохирургии, 1957. Т.1. - С.329-339.
. Благовещенская Н.С.
Отоневрологические симптомы и синдромы. М.: Медицина. 1990. - 430 с.
. Болдырева
Г.Н. Межцентральные отношения в коре головного мозга человека в норме и при
очаговых поражениях диэнцефальных структур. Автореф.дис....доктбиол.наук. М.:
ИВНД и НФ АН СССР, 1978, 28 с.
. Болдырева
Г.Н. Электрическая активность мозга человека при поражении диэнцефальных и
лимбических структур. - М.: Наука, МАИК "Наука/Интерпериодика", 2000.
- 181 с., ил.
. Болдырева
Г.Н. Роль гиппокампальных структур мозга человека в формировании полушарной
специфичности организации ЭЭГ. Тез докл на конф. «Современное состояние
асимметрии мозга». М.: РАН. 2000. - С.
12-14.
. Болдырева
Г.Н., Брагина Н.Н., Доброхотова Т.А. Клинико-электрофизиологическое
исследование больных с поражением гиппокампо-миндалевидного комплекса //
Журн.невропатол. и психиатр. им.С.С.Корсакова. 1972. Т.72. - С.521-530.
. Болдырева
Г.Н., Добронравова И.С. Шарова Е.В., Жаворонкова Л.А Отражение адаптивных
перестроек мозга человека при нарушении церебральных функций в параметрах
межполушарной асимметрии когерентности ЭЭГ
// Журн.высш.нерв.деят. 1993. Т.43.№4. - С.721-729.
. Болдырева
Г.Н., Брагина Н.Н., Маргишвили Г.М., Машерова Е.Л.
Влияние
очага стационарного возбуждения в лимбических структурах
на
изменения пространственно-временной организации ЭЭГ человека
//
Физиология человека. Т.21, 1995. - С.18-27.
44. Болдырева
Г.Н., Шарова Е.В., Добронравова И.С. Роль регуляторных
структур
мозга в формировании ЭЭГ человека // Физиол.челов., 2000.
Т.26.
№4. -С.391-401.
. Болдырева
Г.Н., Жаворонкова Л.А., Шарова Е.В., Добронравова И.С.
Межцентральные
отношения ЭЭГ как отражение системной организации
мозга человека в норме и патологии // Журн.высш.нерв.деят.
2003. Т.53.№6. - С.1093-1099.
46. Большая
медицинская энциклопедия. М.:Советская энциклопедия.
Т.13,
1980. - С.762.
47. Борисова
Е.В. Электрофизиологический и нейрохимический анализ
нейронов
орбитальной области коры и ретикулярной формации среднего
мозга кролика. Автореф...канд.биол.наук. М., 1983, 25 с.
48. Брагина
Н.Н. Клинические поражения гиппокампа и пограничных с
ним
областей. Автореф.дисс. докт.мед.наук.
М. 1966. - 32 с.
. Брагина Н.Н. Особый вид
защитно-рефлекторных реакций при очаговом
поражении гиппокамповой формации // Журн.неврол. и
психиатр.
Т.67 №3, 1967 - С.327-332.
50. Брагина
Н.Н. Клинические синдромы поражения гиппокампа.- М.:
Медицина,
1974. - 216с.
51. Брагина
Н.Н., Доброхотова Т.А. Функциональные асимметрии человека.
М.: Медицина, 1981. 288с.
52. Брагина
Н.Н., Доброхотова Т.А. Функциональная асимметрия мозга и
психические
возможности человека // Взаимодействие полушарий мозга.
Тб. 1982. - С.119-120.
53. Брагина
Н.Н., Доброхотова Т.А. Функциональные асимметрии человека.
М.: Медицина, 1988. 221 с.
54. Брагина
Н.Н., Доброхотова Т.А., Глазман Л.Ю., Ильичева Р.Ф.,
Потапов
А.А. Корсаковский синдром в клинике черепно-мозговой
травмы
// Журн.неврол. и психиатр. Т.88, №5, 1988. - С.18-22.
55. Брагина
Н.Н., Амчеславский В.Г., Баркалая Д.Б., Воронина И.А.,
Хухлаева
Е. А. Ранний послеоперационный период у больных с опухолями
задней черепной ямки // Журн.неврол. и психиатр. Т.97,
№2,
1997. - С.10-14.
56. Бражник
Е.С., Виноградова О.С. Активность нейронов гиппокампа,
лишенного
восходящих стволовых связей // Журн.высш.нерв.деят.
Т.33.
№1, 1983. - С.121-128.
. Брейже
А.Б. Электрофизиологическое изучение зрительного бугра и гиппокампа у человека
// Физиол.журн. Т.53. №9, 1967. - С.1026-1033.
. Бродал
А. Ретикулярная формация мозгового ствола. - М.: Медгиз, 1960.- 99 с.
. Бродал А., Вальберг Ф., Помпеано
О. Вестибулярные ядра. Связи, анатомия,
функциональные корреляции. М.: Л.: Наука, 1966. - 171 с.
60. Буреш
Я., Бурешова О., Хьюстон Д.П. Методики и основные эксперименты по изучению
мозга и поведения. М.: Высшая школа, 1991.
299с.
. Вальдман А.В. Принципиальные
проблемы изучения эмоционального поведения в эксперименте на животных. В кн.:
Экспериментальная нейрофизиология эмоций. Л.: Наука, 1972а. - С. 6-26.
62. Вальдман
А.В. Роль гипоталамуса в эмоционально-поведенческих актах. В кн.: Экспериментальная
нейрофизиология эмоций. Л.: Наука, 1972б. - С.108-123
. Вальдман
А.В. Нейрохимические основы психотропного эффекта //
Фармакология
нейропептидов. М.: Медицина, 1982.
. Вальдман А.В., Звартау Э.Э.,
Козловская М.М. Психофармакология эмоций. М.: Медицина. 1976. - 328 с.
. Вальдман А.В., Цирлин В.А.
Вегетативные механизмы ствола мозга. В кн.: Физиология вегетативной
нервной системы (Серия: Руководство по физиологии...). Л.:Наука, 1981. -
С.341-371.
66. Валюх
Т.П. Электрофизиологический анализ роли гиппокампальных структур в
условнорефлекторной деятельности кроликов. В кн.: ГГГ конференция молодых
физиологов Закавказья. Ереван, 1981. - С.38-44.
. Ванециан
Г.Л. Функциональная асимметрия миндалин кошки при условнорефлекторной
деятельности // Мозг и поведение. М.: Наука, 1990. - С. 69-81.
. Ванециан
Г.Л. Двустороннее взаимодействие гипоталамуса, миндалины и неокортекса на
разных стадиях выработки условного рефлекса // Журн.высш.нерв.деят. Т.43. №5,
1993. - С.889-899.
69. Ванециан Г.Л., Павлова И.В.
Функциональная асимметрия фронтальной коры и латерального гипоталамуса кошек
при инструментальном пищевом условном рефлексе // Журн.высш.нерв.деят. Т.5 3.
№3, 2003. - С.313-320.
70. Вартанян
Г.А., Клементьев В.И. Химическая симметрия и асимметрия
мозга.
Л.:Наука, 1991. - 150с.
71. Вартанян
И.А. Компенсаторно-приспособительные реакции сенсорных систем. В кн.: Основы
физиологии человека, т.3 (ред. Б.И. Ткаченко),
М.:Литера, 1998.- С.91-121.
. Василевская
Н.Е. О структуре и функции висцерохимического анализатора.
- Л., 1971.
73. Ведяев
Ф.П. Модели эмоционально-стрессовых состояний и их
висцеральные
проявления. Материалы симпозиума «Эмоции и висцеральные
функции». Баку: ЭММ, 1974. -
С.11-17.
74. Вейн
А.М., Соловьева А.Д. Лимбико-ретикулярный комплекс и
вегетативная
регуляция. М., Наука, 1973. 268с.
75. Вейн
А.М., Соловьева А.Д. Патологические вегетативные синдромы
(клинико-физиологическая
характеристика). В кн.: Физиология вегетативной
нервной системы. Л.:Наука, 1981. - С.668-710.
76. Викторов И.В. Стволовые клетки
мозга млекопитающих: биология стволовых
клеток in
vivo и in
vitro // Изв. АН. Серия
биолог., 2001. №6. - С.646-655.
. Виноградова
О.С. Гиппокамп и память.- М.: Наука, 1975.- 332 с.
. Виноградова
О.С., Бражник Е.С., Кичигина В.Ф., Стафехина В.С. Модуляция холинергическими
веществами реакции нейронов гиппокампа на сенсорные раздражители //
Журн.высш.нерв.деят. Т.45. №1, 1995. - С.118-130.
. Выготский Л.С. Психология и учение
о локализации психических функций // Собр.соч.: В 6 т. М.: Педагогика, 1982.
Т.6.
. Воронин
Л.Г., Соколов Е.Н. Корковые механизмы ориентировочного рефлекса. - В кн.:
Электроэнцефалографические исследования ВНД. М.:АН СССР, 1962. - 312 с.
. Воронов
В.Г. Выявление статистически значимых особенностей в частотных спектрах
электроэнцефалограмм и приложение этих методов к клинико-диагностическим
задачам // Труды 8-ой Международной конференции «Новые научные технологии в
медицине и экологии». Украина, Гурзуф, 2000, - С.244-245.
. Висцеральная
патология при поражениях центральной нервной системы. Под ред. В.М.Угрюмова,
Л., Медицина, 1975, 304 с.
. Гамбарян
Л.С., Коваль И.Н. Гиппокамп. Ер.: АН Арм. ССР, 1973 -103 с.
. Гасанов Г.Г. Эмоции, висцеральные
функции и лимбическая система. В кн.:
Эмоции и висцеральные функции. Материалы симпозиума.
Баку:
ЭММ, 1974. - С.17-22.
85. Гасанов
Г.Г., Меликов Э.М. Нейрохимические механизмы гиппокампа,
тета-ритм и поведение. М.: Наука, 1986. - 184с.
. Геодакян В.А. Асинхронная
асимметрия // Журн.высш.нерв.деят. Т.43. №3, 1993. -
С.543-561.
87. Гецова
В.М., Йорк Р., Ветцель В. О механизмах участия серотонина в
консолидации
временных связей // Журн. высш. нерв.деят. Т.30. №5,
1980.
- С.988 -993.
. Гильман
И.М., Пилипович А.С., Равикович М.А., Советов А.Н. О компенсаторных
возможностях симметричных областей доминантного и субдоминантного полушарий
головного мозга. Журн.высш.нерв.деят. т.27, №1, 1977.- С.88-97.
. Гильман
И.М., Советов А.Н. Межполушарная асимметрия поздних компонентов вызванных
потенциалов как выражение компенсаторных процессов. Физиол.человека. т.6, №6,
1980.- С.1058-1065.
. Гистология, цитология,
эмбриология: Учебник. / Афанасьев Ю.И. ред. М.:Медицина.
2004. - 766с.
91. Голдберг
Э. Управляющий мозг: Лобные доли, лидерство и цивилизация.
- М.: Смысл, 2003. - 335 с.
92. Гращенков
Н.И. Гипоталамус. Его роль в физиологии и патологии.
М.:
1964, 364 с.
. Гриндель
О.М. Оптимальный уровень когерентности ЭЭГ и его значение //
Журн.высш.нерв.деят. 1980. Т30. №1. - С.62-70.
. Гриндель
О.М., Гершман С.Г., Болдырева Г.Н., Вакар Е.М., Малина З.А., Добронравова И.С.,
Галкина Н.С. Межцентральные отношения в коре больших полушарий по данным
спектра когерентности и фазового спектра ЭЭГ // Журн.высш нерв.деят. 1973.
Т.23, №4. -С.771-781.
. Гриндель
О.М, Брагина Н.Н., Балабанова В.А., Шарова Е.В., Вихерт Т.М. Изменения электрических
процессов в коре больших полушарий человека при очаговых поражениях
гипоталамуса и ствола мозга // Электрофизиологическое исследование стационарной
активноти в головном мозге. М.: Наука, 1983. - С.60-67.
. Гуляева
Н.В. Основы адаптологии. М., 1997. - 108 с.
. Гусельников В.И. Электрофизиология
головного мозга (курс лекций). М.: Высшая школа, 1975. - 423 с.
. Данилова
Н.Н., Крылова А.Л. Физиология высшей нервной деятельности: Учебн.для вузов.
Ростов-на-Дону. 2002. - 479 с.
. Добронравова
И.С. Значение межполушарной асимметрии в развитии компенсаторных процессов
мозга человека // Журн.высш.нерв.деят. 1989. Т39. №5. - С. 819-827.
100.
Доброхотова
Т.А. Эмоциональная патология при очаговых поражениях мозга. - М.:Медицина.
1974. - ....с.
101.
Доброхотова
Т.А., Брагина Н.Н. Функциональная асимметрия и психопатология очаговых
поражений мозга. М.: Медицина, 1977. 359 с.
102.
Доброхотова
Т.А., Брагина Н.Н. Асимметричный мозг -асимметричное сознание //
Журн.высш.нерв.деят. 1993. Т43. №2. - С. 256-261.
103.
Доброхотова
Т.А., Гриндель О.М., Брагина Н.Н.,.Потапов А.А., Шарова Е.В., Князева Н.А.
Восстановление сознания после длительной комы у больных с тяжелой
черепно-мозговой травмой // Журн. невропатологии ипсихиатрии. 1985. Т.85. №6. -
С.720-726.
104.
Дубикайтис
Ю.В., Полякова В.Б. Биоэлектрическая активность головного мозга у больных с
черепно-мозговой травмой при ирритации мозговых структур // Вопросы
нейрохирургии, 1983, №3. -С.14-18.
105.
Дупин
А.М., Барсков И.В., Викторов И.В., Ерин А.Н. Уровень перекисного окисления
липидов в очаге компрессионной ишемии коры головного мозга крыс //
Бюлл.эксперим.биол. и мед. Т.118. №12, 1994.- С.
106.
Жаворонкова
Л.А. Особенности динамики межполушарных соотношений ЭЭГ в процессе
восстановления нервно-психической деятельности человека. Журн.высш.нерв.деят.
Т.40, №2, 1990. -С.238-245.
107. Жаворонкова Л.А. Межполушарные
соотношения электрических процессов мозга здоровых правшей и левшей и больных с
церебральными поражениями. Автореф.дисс...докт.биол.наук. М.,
1999.
. Жаворонкова Л.А.,
Добронравова И.С. Специфика восстановительных процессов мозга у больных с
диэнцефальным и полушарным поражением (когерентный анализ ЭЭГ).
Журн.высш.нерв.деят.
Т.43, №4, 1993. - С.748-757.
109.
Жирмунская
Е.А. Функциональная взаимосвязь больших полушарий мозга человека. Л.: Наука,
1989. - 130с.
110.
Зайченко
М.И. Михайлова Н.Г., Райгородский Ю.В. Активность нейронов префронтальной коры
мозга крыс с разными типологическими особенностями при эмоциональных
воздействиях //. Журн.высш.нерв.деят.
Т.50, №3, 2000. - С. 492-499.
111.
Замбржицкий
И.А. Лимбическая область большого мозга. - М.: Медицина, 1972. - 280 с.
112.
Зверева
З.Ф., Погожева Е.Н., Синяков В.С., Советов А.Н. Межполушарная асимметрия при
одностороннем повреждении мозга. Патолог.физиол.и тер. №2, 1996. - С.6-9.
113.
Зверева
З.Ф., Советов А.Н., Борзунов А.Н. Состояние межполушарной асимметрии в передних
и задних отделах мозга человека в норме и при одностороннем повреждении коры.
Патолог. физиол. и тер. №4, 1998. - С. 3-6.
114.
Зенков
Л.Р., Ронкин М.А. Функциональная диагностика нервных болезней. - М.: Медицина,
1991.- 640с.
115.
Зилов
В.Г. Лимбическая система и ее роль в функциональной системе пищевого и
питьевого поведения. В сб.: Физиология человека и животных. Т.20. М., 1977,
С.5-57.
116.
Зилов
В.Г. Химическая интеграция в механизмах пластичности мозга.
Вестн.Росс.Акад.мед.наук. 1997, №12. - С.50-52.
117.
Зилов
В.Г., Рогачева С.К. Влияние электрической стимуляции различных областей
головного мозга на пищевое поведение кроликов // Журн.высш.нерв.деят. Т.25, №5,
1975. - С. 1076-1082.
118.
Зимкина
А.М. Ретикулярная формация и ее роль в регуляции функций мозга в норме и
патологии. Физиол.журн.Т. 44, №4, 1958.-С. 369.
119.
Зимкина
А.М. О критериях устойчивости в проявлениях деятельности центральной нервной
системы человека. Физиол.журн.Т.58, №7, 1972. С.1011- 1018.
120.
Зимкина
А.М. О компенсации нарушенных функций центральной нервной системы. Физиол.журн.
Т.68, N 7, 1982, С. 918-925.
121.
Зухарь
А.В. Типы высшей нервной деятельности, стрессоустойчивость и репродуктивная
функция. // Дисс.канд.биол.наук. М.: ИВнД
и НФ РАН, 1985. - 155 с.
122.
Егоров
Ю.В., Кузнецова Г.Д. Форма электрических полей при поляризации мозговой ткани
// Электрическая активность головного мозга при образовании простых форм
временной связи. М.: Наука, 1972. 99 с.
123.
Ильюченок
Р.И. Нейро-гуморальные механизмы ретикулярной формации ствола мозга. - М.:
Наука, 1965. - 258 с.
124.
Ильюченок
Р.Ю., Гилинский М.А. Конструкция и медиаторы ретикуло-корковых связей. - Л.:
Наука, 1971. - 152 с.
125.
Иоффе
М.Е. О механизмах двигательного обучения. Журн.высш.нерв.деят. Т.41, №5. 1991.
- С.907-925.
126.
Иоффе
М.Е., Плетнева Е.В., Сташкевич И.С. Природа функциональной моторной асимметрии
у животных: состояние проблемы. Журн.высш.нерв.деят. Т.52, №1, 2002. - С.5-16
127.
Карамян
а.И. Эволюция функций мозжечка и больших полушарий головного мозга.
М.-Л.:Медгиз, 1956.
128.
Карамян
А.И., Соллертинская Т.Н. Роль структур лимбического мозга в регуляции
поведенческой деятельности в филогенезе позвоночных. В кн.: Психофармакология
эмоций. М.: Медицина, 1976.
-
С. 32-58.
129. Карамян А.И., Соллертинская
Т.Н. Значение структур лимбического мозга в условнорефлекторной деятельности у
кроликов // Физиол.журн. Т.68, N 7, 1982, С. 868-884.
130.
Карцева
А.Г. Морфо-функциональная организация связей мозговых структур, участвующих в
регуляции кровообращения. Автореф. дисс. докт.мед.наук.
Киев, 1985. 37 с.
131.
Квирквелия
Л.Р. Межгиппокампальные и внутригиппокампальные функциональные взаимодействия
// Автореф.дис. докт.биол.наук. М.:ИВНД и НФ АН СССР, 1987. 38 с.
132.
Кичигина
В.Ф., Кудина Т.А. Сенсорные реакции нейронов гиппокампа кролика при
функциональном выключении структур, управляющих тета-ритмом.
Журн.высш.нерв.деят. Т.51, №2, 2001. - С.228-235.
133.
Коган
А.Б. Вероятностно-статистический принцип нейронной организации функциональных
систем мозга. Докл. АН СССР, 1964, т.154, с.1231-1240.
134.
Коваль
И.Н., Саркисов Г.Т. Условные двигательные рефлексы после повреждения гиппокампа
и свода у кошек. В сб.: Психофизиология эмоций. М.: Медицина, 1976. -
С.121-130.
135.
Коваль
И.Н., Саркисов Г.Т., Гамбарян Л.С. Септо-гиппокампальная система и организация
поведения.- Ер.: изд.АН Арм.ССР, 1986- 127 с.
136.
Козловская
М.М., Вальдман А.В. Вегетативные корреляты эмоциональных реакций. В кн.:
Экспериментальная физиология эмоций. Л.: Наука, 1972. - С. 173-210.
137.
Корочкин
Л.И. Стволовые клетки - один из путей регенерации нервной ткани. Изв. АН. Серия
биол., 2001, №6.- С.666-671.
138.
Котляр
Б.И., Шульговский В.В. Физиология центральной нервной системы. М.: МГУ, 1979. -
349 с.
139.
Кратин
Ю.Г., Сотниченко Т.С. Неспецифические системы мозга. -Л.: Наука, 1987.- 159 с.
140.
Кругликов
Р.И. Нейрохимические механизмы обучения и памяти. М.:
Наука,
1981. - 211с.
141.
Крылова
Н.В., Искренко И.А. Анатомия в схемах и рисунках (спинной, головной мозг и
черепные нервы): Учеб.пособие.- М.: Изд-во УДН, 1986. - 168 с.
142.
Крыжановский
Г.Н. Детерминантные структуры в патологии нервной системы. - М.: Медицина,
1980. - 360 с.
143.
Кураев
Г.А., Сороколетова Л.Г. Компенсация нарушенных функций в нервной системе.
Ростов-Дон, 1996. - 111 с.
144.
Лагутина
Н.И., Хасабов Г.А., Урманчеева Т.Г. О взаимосвязи гиппокампа и стриопаллидума в
организации сложного поведения. В кн.: Психофармакология эмоций. М.: Медицина,
1976. - С.109-114.
145.
Лапина
И.А., Яичников И.К., Щабанов П.Д., Бородкин Н.С. Межполушарные взаимоотношения
структур мозга при разных функциональных состояниях центральной нервной системы
// Физиол.ж.СССР, 1984. Т.70, №1. - С.3-8. 147.
Левшина
И.П., Гехт К., Нгуен Ван-Хай. К значению одностороннего и двустороннего
повреждения гиппокампа в регуляторных процессах центральной нервной системы //
Журн.высш.нерв.деят. 1977. Т.27. №3.- С.366-368.
148.
Левшина
И.П., Л.В.Ноздрачева, Шуйкин Н.Н. Особенности внешнего дыхания как показатель
активности клеточных дыхательных ферментов мозга крыс // Бюлл.эксп.биол. и мед.
1997. Т.124, №8. - С.138-140.
149.
Леонтович
Т.А. К проблеме эмоций // Усп.соврем.биол., 1968, 85,
150. Леонтович Т.А. Компенсаторные
изменения дендритов клеток взрослого мозга в экспериментах на млекопитающих и в
патологии мозга человека. Восстановительная неврология: Тез докл. 5-го
выезд.засед.эксперт.сов. по молекул.биол. и мед.биотехнол. АМН СССР, науч. сов.
АН и АМН СССР по физиол. человека и президиумап Вост.-Сиб. Фил. СО АМН СССР,
посв.памяти В.М. Бехтерева.
Иркутск,
1990. С. 49-50.
. Ливанов М.Н.
Пространственно-временная организация потенциалов в деятельности головного
мозга. Избранные труды. М.: Наука, 1989,
398 с.
152.
Ливанов
М.Н., Думенко В.Н. Нейрофизиологический аспект исследований системной
организации деятельности головного мозга // Усп.физиол.наук. 1987. Т.18. №3. -
С.6-16.
153.
Лиманский
Ю.П. Рефлексы ствола головного мозга. Киев: Наук.думка, 1987.- 240с.
154.
Лосева
И.В., Силаков В.Л. Влияние ликвора животных с повреждением моторной коры на
компенсацию двигательных нарушений. Физиол.журн. т.73, №12, 1987.- С.1608-1614.
155.
Лурия
А.Р. Основы нейропсихологии. Учеб. Пособие для студ. Высш. Учеб. Заведений. -
М.: Издательский центр «Академия», 2002. - 384 с.
156.
Лурия
А.Р, Смерницкая Е.Г. // Нейропсихология. 1977. Т.15, №1. -
С.175-178.
157. Магаева С.В., Морозов С.Г.
Нейроиммунофизиология. М., 2005. - 158с.
158.
Магнус
Р. Установка тела. - М.Л.: Изд. АН СССР, 1962.- 624 с.
159.
Мадорский
С.В. Эмоциональные нарушения при поражении структур височной доли мозга.
М.:Наука, 1985.- 152с.
160. Майорчик В.Е. Опухоли в области
ЗЧЯ. В кн.: Клиническая электроэнцефалография под ред.В.С.Русинова. М.:
Медицина, 1973. - С.173-146.
161.
Макаров
В.А. Лимбическая система и половые функции. В сб.: Физиология человека и
животных. Т.20. Физиология лимбической системы. М., 1977.- С.66-107.
162.
Макаренко
А.Н. Нейрофизиологические и нейрохимические аспекты протекторного влияния
неокортекса при воздействии и передозировке общих анестетиков. Автореф.дисс.
докт.мед.наук. -М.:ИВНД и НФ РАН, 1993. - 51с.
163.
Матинян
Л.А.Эволюционные аспекты компенсации функций поврежденного спинного мозга //
Журн.высш.нерв.деят, 2003. Т.53, №3. - С.279-289.
164.
Меликов
Э.М. Роль гиппокампа и его моноаминореактивных систем в механизмах сна кошки //
Журн.высш.нерв.деят. Т.37. №2, 1987. - С.246-253.
165.
Мельников
А.В., Новикова М.Р., Куликов М.А., Шарова Е.В., Сидоров Б. М. Корреляционная
зависимость выбора направления движения крыс от других показателей в тесте
открытого поля, вынужденного плавания и теста Кинга. Мат.конф. «Актуальные
вопр.функц.межполуш.асимметрии». М. 2001. - С.106-110.
166.
Микляева
Е.И., Иоффе М.Е., Куликов М.А. Предпочтение одной конечности у крыс - результат
обучения в эксперименте или индивидуальная особенность? Журн.высш.нерв.деят.
Т.38, №5, 1988. - С.881-888.
167.
Микляева
Е.И. Моторная асимметрия при выработке локальных инструментальных рефлексов у
белых крыс. Дисс.на соиск..канд.наук. М., 1989.
168.
Михайлова
Н.Г. Участие эмоциогенных механизмов гипоталамуса в структурно-функциональной
организации поведенческого акта. Дисс.дмн. М., 1994. - 231с.
169.
Михайлова
Н.Г., Зайченко М.И. Нейроны правой и левой префронталдьной области коры мозга
крыс при стимуляции эмоциогенных зон // Журн.высш.нерв. деят. 1998. Т.48. №3.-
С.431-437.
170.
Молодцова
Г.Ф. Половые и межполушарные различия в вовлечении серотонина гиппокампа и
миндалевидного комплекса в обработку новой и повторно предъявляемой информации
у крыс // Журн.высш.нерв.деят. Т.49, №3, 1999. - С.408-415.
171.
Молодцова
Г.Ф. Влияние факторов пола и полушарной локализации на вовлечение серотонина
фронтальной коры, стриатума и прилежащего ядра в обработку новой и повторно
предъявляемой информации у крыс. Журн.высш.нерв.деят. Т.51, №1, 2001. -
С.56-60.
172.
Морфология
человека: Учеб. Пособие.-2-у изд., перераб., доп./Под ред. Никитюка Б. А., Чтецова
В.П. - М.: Изд-во МГУ, 1990.- 344 с.
173.
Мосидзе
В.М., Рижинашвили Р.С., Самадашвили З.В., Турашвили Р.И. Функциональная
асимметрия мозга. Тб.:Мецниереба, 1977. - 120 с.
174.
Мотлох
Н.Н. Компенсаторный резерв и межуровневые соотношения при регенерации;
Научн.центр биол. Исслед. АН СССР. Пущино. 1987. - 150 с. ил. Библиогр. №83
назв. Рус.Рукопись деп. В ВИНИТИ 22.07.87. № 5290-В87.
175.
Моторина
М.В. Исследование гипоталамо-кортикальных связей у кролика //
Журн.эволюц.биохим и физиол. Т.4, №2, 1968. - С.187-193.
176.
Мурик
З.С. Межполушарная асимметрия и типологические особенности нервной системы
крыс. Журн.высш.нерв.деят. Т.40, № 5, 1990. - С.963-967.
177.
Мэгун
Г. Бодрствующий мозг. - М.: издательство Иностранной литературы, 1961. - 126 с.
178.
Наливаева
Н.Н., Плеснева С.А., Ковешникова Т.Г., Васильева Ю.В., Варлинская Е.И.,
Клементьев Б.И. Сравнительно-биохимический анализ неокортекса крыс-правшей,
левшей и амбидекстров // Докл. АН, 1993. Т.328. №1. - С.116-118.
179.
Наута
У. Некоторые связи лимбической системы // Механизмы целого мозга. М., 1963. -
С.182-198.
180.
Наута
У.Дж., Кейперс Г.Г. Некоторые восходящие пути ретикулярной формации. В сб.:
Ретикулярная формация мозга. М.: Гос.изд.мед.лит., 1962. - С.13-37.
181.
Незлина
Н.И. Нарушение и восстановление двигательных функций у собак после половинной
перерезки мозгового ствола. Дисс.... канд. биол. наук. М. 1957. - 180 с.
182.
Нейрохимия.
Ред.Проханова М.И. - Л.: ЛГУ, 1979. - 271 с.
183.
Ноздрачев
А.Д., Чернышова М.П. Висцеральные рефлексы. Л.: ЛГУ, 1989. - 168с.
184.
Ноздрачев
А.Д., Поляков Е.Л. Анатомия крысы (Лабораторные животные)/ Под ред.
А.Д.Ноздрачева. СПб.: Лань, 2001.- 464 с.
185.
Нуцубидзе
М.А. Эмоциональная и замыкательная функция лимбической системы. - Тб.:
Мецниереба, 1969. -186 с.
186.
Ониани
Т.Н. Интегративная функция лимбической системы. - Тб.:
Мецниереба,
1980. - 302 с.
187. Опитц Б., Саркисова К.Ю.
Межполушарная асимметрия перекисного окисления липидов мозга крыс с разным
типом поведения как прогностический показатель их устойчивости к церебральной
ишемии и эффективности противоишемического действия субстанции Р. Докл.АН.
Т.346, №2, 1996. - С.275-277.
188.
Основы
физиологии человека. Клинико-физиологические аспекты. Учебник под ред. Ткаченко
Б.И. - М.: Литера, 1998, с.473.
189.
Отмахов
Н.А. Нейрональная сеть гиппокампа: морфологический анализ // Успехи
физиол.наук. Т.24. №4, 1993. - С.79-101.
190.
Павлов
И.П. Собрание трудов. М.: Наука, 1951, Т.3.
191.
Павлова
И.В. Функциональная асимметрия медиального гипоталамуса кролика при реакции
избегания // Журн.высш.нерв.деят. 1998. Т.48. №3.- С.396-404.
192.
Павлова
И.В. Функциональная асимметрия мозга при мотивационных
и
эмоциональных
состояниях.
Дисс....докт.биол.наук. М. 2001. -225 с.
193.
Павлыгина
Р.А., Соколов С.С. Роль доминанты в целенаправленном поведении животных. В кн.:
Электрофизиологическое исследование стационарной активности в головном мозге.
М.: Наука, 1983. - С.34-54.
194.
Пейпец
Дж.Б. Висцеральный мозг, его строение и связи. В сб.: Ретикулярная формация
мозга. М.: Гос.изд.мед.лит., 1962. - С. 520532.
195.
Пигарева
М.Л. Лимбические механизмы переключения. М.: Наука, 1978. - 151 с.
196.
Пигарева
М.Л. Экспериментальное исследование нейропсихологии эмоций. Дисс.
докт.биол.наук. М., 1983. - 401 с.
197.
Пигарева
М.Л. Формирование пищевого условного рефлекса при вероятностном подкреплении у
крыс с повреждением фронтальной коры // Журн.высш.нерв.деят. 1984. Т.34. №6.-
С.1100-1108.
198.
Плицитый
Д.Ф., Магаева С.В. О регуляции иммуногенеза структурами гиппокампа // Доклады
АН СССР. 1970. Т.194. №1. - С.232-234.
199. Плотников М.Б, Ваизова О.Е.,
Суслов Н.И. Анализ изменений спектра мощности электроэнцефалограммы на новой
модели ишемии мозга у крыс// Бюлл.эксперим.биол.и мед. Т.118, №12, 1994. -
С.565-567.
200.
Подачин
В.П., Мусалов Г.Г., Незлина Н.И. Структурно-функциональные основы компенсации
функций при травме спинного мозга. М.: Наука, 1983, 190 с.
201.
Подачин
В.П., Сидоров Б.М. Компенсаторные процессы при повреждении лимбической системы.
М.: Наука, 1988. - 160 с.
202.
Подачин
В.П. Проблема компенсации нарушенных функций в научной деятельности Э.
А.Асратяна и современное состояние проблемы. Тез.докл.конф. «Восстановл.функций
повр.мозга» М.: ВНД и НФ РАН, 1977.
- С.2.
203. Полякова В.Б. Роль состояния
ствола головного мозга при определении степени компенсации механизмов регуляции
вегетативных функций (на примере черепно-мозговой травмы) // Журн.неврол. и
психиатр. Т.99, №8, 1999. - С.9-11.
204.
Пошивалов
В.П. Экспериментальная психофармакология агрессивного поведения. Л.:Наука,
1986. - 175с.
205.
Преображенская
Л.А. Функциональная асимметрия электрической активности неокортекса в процессе
выработки условного пищевого рефлекса у собак // Журн.высш.нерв.деят. Т.50. №3,
2000. - С.434-446.
206. Раева С.Н., Брагина Н.Н.,
Фалер Т.О. Особенности электроэнцефалограммы, связанные с поражением разных
отделов ствола мозга у человека // Журн.высш.нерв.деят. Т.16, №5, 1966. -
С.895-901.
207.
Романова
Л.К. Дыхательная система. В сб.: Структурные основы адаптации и компенсации
нарушенных функций. Ред. Д.С.Саркисов. М.:Медицина, 1987. - 448с.
208.
Романова
Г.А., Барсков П.В., Советов А.Н., Викторов И.В. Нарушение интегративной
деятельности мозга крысы при двусторонней фокальной компрессионной ишемии
лобной коры // Бюлл.эксперим.биол.и мед. Т.118, №12, 1994.-С.568-571.
209.
Романова
Г.А., Векшина Н.Л., Советов А.Н. Участие моноаминов в восстановительных
реакциях центральной нервной системы при экспериментальном повреждении передних
отделов коры головного мозга. Бюлл.экспер.биол. и мед. Т.90, №11, 1980.-
С.536-538.
210.
Росси
Дж.Ф., Цанкетти А. Ретикулярная формация ствола мозга. -М.: издательство
Иностранной литературы, 1960. -263 с.
211.
Русалова
М.Н., Костюнина М.Б. Асимметрия биоэлектрической активности мозга при
переживании негативных эмоций // Доклады АН. Т.380, №5, 2001. - С.701-703.
212.
Русинов
В.С., Гриндель О.М., Болдырева Г.Н. Исследование динамики межцентральных
отношений в коре больших полушарий головного мозга методом спектрального
анализа ЭЭГ // Механизмы деятельности головного мозга. Тбилиси: Мецниереба,
1975. - С.365-374.
213.
Русинов
В.С., Гриндель О.М. Отражение состояния и функций мозга человека в структурах
межцентральных отношений по данным спектрально-корреляционного анализа ЭЭГ //
Усп.физиол.наук. 1987. Т.18. №3. - С.39-51.
214.
Русинов
В.С., Гриндель О.М., Болдырева Г.Н., Вакар Е.М. Биопотенциалы мозга человека.
Математический анализ. М.:Медицина. 1987. - 254 с.
215.
Русинова
Е.В. Влияние стимуляции поля СА3 дорзального гиппокампа на двигательную
поляризационную доминанту у кроликов // Журн.высш.нерв.деят. Т.39, №2, 1989. -
С.301-308.
216.
Русинова
Е.В. Корково-подкорковые взаимоотношения при формировании временной связи на
модели двигательной поляризационной доминанты. Автореф...докт.биол.наук. М.
1996. - 31 с.
217.
Рябинская
Е.А., Валуйская Т.С., Ашихмина О.В. Пространственно-моторная асимметрия в
поведении крыс. Журн.высш.нерв.деят. Т.34, №6, 1984. - С.1078-1085.
218.
Савельев
С.В., Негашева М.А. Практикум по анатомии мозга человека: Учебное пособие для
студентов вузов.- М.: ВЕДИ, 2001.192 с.
219.
Саркисов
С.А. Очерки по структуре и функции мозга. М: Медицина,
1964.- 300с.
220.
Саркисов
Д.С. Общие закономерности компенсаторно-приспособительных реакций и их
структурное обеспечение. Материальные основы надежности биологических систем. В
сб.: Структурные основы адаптации и компенсации нарушенных функций. Ред.
Д.С.Саркисов. М.:Медицина, 1987. - 448с.
221.
Саркисова
К.Ю., Куликов М.А., Ноздрачева Л.В., Коломийцева И.А. Значение
индивидуально-типологических особенностей поведения
и
энергетического метаболизма мозга при прогнозировании течения экспериментальной
патологии // Нарушения высшей нервной деятельности, их патогенез и
нейропептидная коррекция. М.:. 1992. с.26.
222.
Саркисова
К.Ю., Куликов М.А. Новая экспериментальная модель депрессии: WAG/Rij
// Докл.АН. 2000. Т. 344. №5. - С. 706-710.
223.
Сепп
Е.К., Цукер М.Б., Шмидт Е.В. Нервные болезни. М.: Медгиз, 1950. - С.172.
224.
Селье
Г. Стресс без дистресса.- М.: Прогресс, 1982 - 124с.
225.
Симонов
П.В. Что такое Эмоция? М.: Наука, 1966.- 94с.
226.
Симонов
П.В.Теория отражения и психофизиология эмоций. М.: Наука, 1970. 142 с.
227.
Симонов
П.В. Эмоциональный мозг М.: Наука, 1981.- 215 с.
228.
Симонов
П.В. Мотивированный мозг М.: Наука, 1987 - 238с.
229.
Симонов
П.В. О нервных центрах эмоций // Журн.высш.нерв.деят. 1993. Т.43. №3. - С.
514-529.
230.
Симонов
П.В. Функциональная асимметрия лимбических структур мозга //
Журн.высш.нерв.деят. Т.49, №1, 1999. - С. 22-27.
231.
Симонов
П.В. Лекции о работе головного мозга: Потребностно-информационная теория высшей
нервной деятельности. - М.: Наука, 2001. - 95 с.
232.
Симонов
П.В., Русалова М.Н., Преображенская Л.А., Ванециан Г. Л. Фактор новизны и
асимметрия деятельности мозга // Журн.высш.нерв.деят. Т.45, №1, 1995. - С.
13-17.
233.
Смирнов
Л.И. Патологическая анатомия и патогенез травматических заболеваний нервной
системы. М.: Медгиз, 1949. Ч.2.
234.
Смирнова
Н.Я. Висцерально-вегетативные нарушения у больных после удаления опухолей
срединно-базальной локализации и методы их патогенетической терапии. Дисс.
соиск. . канд. мед. наук. М., 1986.
235.
Советов
А.Н. Восстановительные и компенсаторные процессы в центральной нервной
системе.- М.: Медицина, 1988.- 144 с.
236.
Советов
А.Н., Гильман И.М. О соотношении электрической активности в симметричных
областях коры мозга при его односторонних поражениях // Журн.высш.нерв.деят.
Т.25, №2, 1975. - С.388-395.
237.
Соллертинская
Т.Н., Шорохов М.В., Цвященко Т.Ю. Нейрогормоны как модуляторы нейропластичности
мозга и роль лимбических структур в механизмах их влияния (эволюционные аспекты
исследования) // Материалы конференции «Актуальные вопросы функц.межполуш.
асимметрии». М. 2005. - С.254-257.
238.
Судаков
К.В. Индивидуальная устойчивость к эмоциональному стрессу. М., 1998. - 267 с.
239.
Сташкевич
И.С., Куликов М.А. К вопросу о формировании латерализованного двигательного
навыка у крыс // Журн.высш.нерв.деят. Т.50, №3, 2000. - С.457-463.
240.
Сташкевич
И.С., Мац В.Н., Куликов М.А. Индивидуальные и популяционные особенности
формирования функциональной моторной асимметрии у крыс. Тез.докл. на конф.
Современное состояние асимметрии мозга. М., 2000. - С. 9-10.
241.
Степаненко
А.Ю., Архипова Н.А., Хухлаева Е.А., Коршунов А.Г. Зависимость результатов
хирургического лечения очаговой эпилепсии от локализации патологического
процесса и эпилептогенной зоны. Тез.докл. на междунар.конф. «Болезнь Паркинсона
- диагностика, лечение, реабилитация» от 30.09.2000. М.
242.
Трауготт
Н.Н. Межполушарные взаимоотношения при локальных поражениях головного мозга. В
сб.: Нейропсихологический анализ межполушарной асимметрии мозга. - М.:Наука,
1986. - С.14-22.
243.
Трофимова
Е.В. Особенности межполушарного взаимодействия у правшей и левшей по данным
когерентного анализа ЭЭГ // Журн.высш.нерв.деят. Т.50, №6, 2000. - С.943-951
244.
Угрюмов
В.М., Теплов С.И., Тиглиев Г.С. Регуляция мозгового кровообращения. - Л.:
Медицина, 1984. - 136 с.
245.
Удалова
Г.П., Михеев В.В. Роль функциональной межполушарной асимметрии в формировании
предпочтения направления движения у крыс // Журн.высш.нерв.деят. Т.32, №4,
1982. - С.633-640.
246.
Умрюхин
А.Е. Серотонин в паравентрикулярном ядре гипоталамуса и дорсальном гиппокампе
при стрессорных нагрузках у крыс с различной двигательной активностью.
Авт.дисс. . канд.мед.наук. М. 2002. - 24 с.
247.
Урганджян
Т.Г. Возрастные особенности компенсаторного восстановления функий. Изд. АН Арм.
ССР, Ереван, 1973
248.
Урганджян
Т.Г. Развитие научного наследия Э.А.Асратяна в области физиологии компенсации
функций центральной нервной системы. В сб.: Механизмы динамической локализации
и компенсации функций центральной нервной системы - Ереван: АН Арм.ССР, 1986.-
С.9-16.
249.
Урманчеева
Т.Г. Значение гиппокампа в корково-подкорковых взаимоотношениях. Авт.дисс. .
докт.мед.наук. М. 1970-49с.
250.
Фокин
М.С. Интенсивная терапия и реанимация в послеоперационном периоде у больных с
опухолями задней черепной ямки. Дисс...канд.мед.наук. М.: ИНХ им.Н.Н. Бурденко,
1985. - 183 с.
251.
Фурдуй
Ф.И. Физиологические механизмы стресса и адаптации при остром действии
стресс-факторов. Кишинев, Штиница, 1986. С.- 304.
252.
Хамильтон
Л.У. Основы анатомии лимбической ситемы крысы. М.: МГУ, 1984. - 200 с.
253.
Хананашвили
М.М. Роль лимбической системы мозга в организации и регуляции эмоций. В кн.:
Экспериментальная нейрофизиология животных. Л.: Наука, 1972. - С. 63-76.
254.
Хананашвили
М.М. Патология высшей нервной деятельности. М.: Медицина, 1983. - 320 с.
255.
Хаспеков
Л.Г., Онуфриев М.В., Викторов И.В., Гуляева Н.В. Действие ишемии и
реоксигинации на накопление нитритов в культурах клеток-зерен мозжечка крыс //
Нейрохимия. Т.17, №4, 2000.- С.267-270.
256.
Хомская
Е.Д. Мозг и активация. М.: МГУ, 1972. - С.16-21.
257.
Хомская
Е.Д. Нейропсихология. М.: МГУ, 1987. - 288 с.
258.
Хомская
Е.Д. Нейропсихология. М.: УМК «Психология», 2002. -416 с.
259.
Хомская
Е.Д., Ефимова И.В., Будыка Е.В., Ениколонова Е.В. Нейропсихология
индивидуальных различий. М.: Рос. педагог агенство, 1997. - 281 с.
260.
Чепурнов
С.А., Чепурнова Н.Е. Миндалевидный комплекс. М.: МГУ, 1981, 256 с.
261.
Чернышев
С.Н. Клиника и хирургическое лечение спастической кривошеи.
Автореф..канд.мед.наук. Л.:ИНХ им.Поленова, 1983. -
11с.
262. Чилингарян Л.И.
Индивидуально-типологические особенности высшей нервной деятельности собак и
межполушарная асимметрия электрической активности гиппокампа и миндалины //
Журн.высш.нерв.деят.2002. Т.52. №1. - С. 47-56.
263.
Чилингарян
Л.И., Богданов Н.Н. Влияние анксиолитиков и анксиогена на электрическую
активность фронтальной коры и лимбических структур собак с разными свойствами
нервной системы // Журн.высш.нерв.деят. 1998. Т.48. №6. - С. 1014-1026.
264.
Шарова
Е.В. Адаптивно-компенсаторные перестройки биоэлектрической активности мозга
человека при повреждении стволовых образований. Дисс.... докт. биол.наук. М.
1999. - 328 с.
265.
Шарова
Е.В., Куликов М.А., Баркалая Д.Б., Амчеславский В.Г. Межцентральные отношения
корковой биоэлектрической активности мозга человека после оперативного
вмешательства на стволовых образованиях // Журн.высш.нерв.деят. 1991. Т.41. №2.
- С.246
266.
Шарова
Е.В., Брагина Н.Н., Баркалая Д.Б., Куликов М.А., Сировский Э. Б. Сопоставление
структуры корковой биоэлектрической активности мозга человека при остро
развивающихся патологических очагах на разных уровнях ствола //
Журн.высш.нерв.деят. 1992. Т.42, №3.- С.439-448.
267.
Шарова
Е.В., Бородкин О.М., Гогитидзе Н.В., Лукьянов В.И., Муханов Т. К.
Функциональное значение показателей пространственно-временной организации ЭЭГ у
больных с черепно-мозговой травмой // Физиол.чел. Т.18, №6. 1992. - С.22-30.
268.
Шарова
Е.В., Болдырева Г.Н., Брагина Н.Н., Куликов М.А. Адаптивно-компенсаторные
перестройки внутриполушарного взаимодействия электрических процессов мозга
человека при поражении стволовых структур // Журн.высш.нерв.деят. 1993. Т.43.
№6.
- С.1089.
269.
Шарова
Е.В., Манелис Н.Г., Куликов М.А., Баркалая Д.Б. Влияние стволовых структур на
формирование функционального состояния больших полушарий головного мозга
человека // Журн.высш.нерв.деят. 1995. Т.45, №5.- С.876-885.
270.
Шахнович
А.Р. Мозг и регуляция движений глаз. М.:Медицина, 1974. - 160с.
271.
Шмидт
Р., Тевс Г. (ред). Физиология человека. Т.1 (нервная система). М.: Мир. 1985. -
265 с.
272.
Шульговский
В.В. Физиология центральной нервной системы. Учебник - М.: МГУ, 1997.- 397 с.
273.
Юринов
А.В. В кн.: Актуальные проблемы патофизиологии. СПб. 2001.- С.203.
274.
Ярославцев
В.Л. (ред.) Восстановительная неврология: Тез.докл.5-го выездного заседания
экспертного совета по молек.биол. и мед.биотехнол. АМН СССР, науч.сов. АН и АМН
СССР по физиологии человека. Иркутск, март, 1990. Иркутск: Гос.мед.ин-т, 1990.
- 260с.
275. Abatzides G.J., Kitsios
A. The role of rehabilitation in the treatment of balance disorders. Review //
J. Back Musculoskeletal Rehabil. 1999;
12(2):
101-112.
276. Abe
K. Modulation of hippocampal long-term potentiation by the amygdale: a synaptic
mechanism linking emotion and memory // Jpn.J.Pharmacol. 2001 May; 86(1):18-22.
277.
Adolphs R. Neural systems for
recognizing emotion // Current Opinion in Neurobiol., 2002. 12(2).
- P.169-177.
278.
Adolphs R., Damasio H., Trancl D.,
Damasio A.R. Cortical systems for the recognition of emotion in facial
expressions // J.Nerosci. 1996. V.16, N23. - P.7678-7687.
279. Alanis
J., Mascher D.J., Miyamoto J. Relationship between self-susteined activity of
the cerebral certex and the efferent discharge of cardic sympathetic and
parasympathetic fibers // Jpn.J.Physiol. 1966, 16.- P.316-334.
280.
Alice C., Paul A.E., Sansom A.J.,
Maclennan K., Darlington C.L., Smith P.F The effects of steroids on vestibular
compensation and vestibular nucleus neuronal activity in the guinea pig.
J.Vestib.Res. v.8, N3, 1998.-P.201-207.
281. Amaral
D.G., Sinnamon H.M. The locus coeruleus: neurobiology of a central norarenergic
nucleus. Progr.in Neurobiol. 1977, V.9.-
P.147-196.
282.
Arnolds D.E., Aitink J.W., Boeyina
P., Lopes da Silva F. EEG hippocampique de home, du chien et du chat //
Rev.electroencephalogr.neurophysiol.clin. V.9. №4, 1979. - P.326-334.
283. Azrin
N.H., Hutchinson R.K., Hake D.E. Elicitation of aggression by physical blow. -
J.Exp.Anal. of Behav., 1965. V.8, №1. - P.55-57.
284. Bacskai
T., Szekely G., Matesz C. Ascending and descending projections of the lateral
vestibular nucleus in the rat // Acta Biol. Hung.
2002; 53 (1-2): 7-21.
285. Basso D.M., Beattie M.S.,
Bresnahan J.C. Descending systems contributing to locomotor recovery after mild
or moderate spinal cord injury in rats: experimental evidence and a review of
literature // Restor Neurol
Neurosci. 2002; 20(5): 189-218.
286. Bland
B.H. The physiology and pharmacology of hippocampal formation theta-rhythms //
Progr.Neurobiol. 1986 (1). - P.1-54.
287. Blonder
L.X., Bowers D., Heilman K.M. The role of thew right hemisphere in emotional
communication // Brain. 1991. V.114,
N3.
-
P.1115-1127.
288. Beauregard M., Gold D.,
Evans A.C., Chertkow H.A role for the hippocampal formation in implicit memory:
a 3-D PET study // Neuroreport.
1998 Jun 1; 9(8):1867-1873.
289.
Burns S.M., Wiss J.M. The
involvement of the anterior cingulated cortex in blood pressure control // Brain
Res. 1985. V. 340. N1. - P.71-77.
290. Carlson
J.N., Glick S.D.Cerebral lateralization as a source of interindividual
differences in behavior // Experientia. - 1989. V.45. №9.
-
P.788-798.
291. Cahoon
D.D. A behavioristic analysis of aggression. The psychol.Res. 1972. V.22, №3. -
P.463- 476.
292.
Cardinal R.N., Parckinson J.A., Hall
J., Everitt B.J. Emotion and motivation: the role of the amygdale, ventral
striatum, and prefrontal cortex // Neurosci. Biobehav. Rev. 2002
May; 26(3): 321-352.
293.
Cass S.P., Goshgarian H.G. Increased
glial fibrillary acidic protein immunoreactivity in astrocytes within the
lateral vestibular nucleus of the cat following labyrinthectomy and vestibular
neurectomy. Ann.Otol.Rhinol.Laryngol. v.99. N3 Pt
1,
1990.- P.221-7.
294.
Chida Y., Toyosawa K. Rhythmic slow
activity (RSA) recorded from the bilateral hippocampus in the freely moving
rats // J.Physiol. (1992) V.446. Cambridge Meeting 18-20
July 1991.
295.
Gion M.L., Chon M. " Barrel
rotation" induced by somatostatin in the non-lesioned rats // Br.Res,
1975. V.96. - P.138-141.
296. Concha
M.L., Wilson S.W. Asymmetry in the epithalamus of vertebrates // J.Anat. 2001,
199 (Pt 1-2): 63-84/
297.
Cransac H., Peyrin L., Farhat F.,
Cottet-Emard J.M., Pequignot J.M., Reber A. Effect of hemilabyrinthectomy on
monoamine metabolism in the medial vestibular nucleus, locus coeruleus, and
other brainstem nuclei of albino and pigmented rats. J.Vestib.Res. v.6, N4,
1996.- P.243-253.
298. Danenberg
V.H. Hemispheric laterality in animals and the effect of early experience. In.:
The behavioral and brain sciences. 1981, 4. -P.1-49.
299. Darlington
C.L., Flohr H., Smith P.F. Molecular mechanisms of braistem plasticity. The
vestibular compensation model. Mol.Neurobiol. v.5, N
2-4, 1991. - 355-368.
300. Darlington
C.L., Smith P.F., Gilchrist D.P. Comparison of the effects of ACTH-(4-10) on
medial vestibular nucleus neuron in braistem slices from labyrinthine-intact
and compensated guinea pigs. Neurosci.Lett. v.145, N1,
1992. - P.97-99.
301. Darlington
C.L., Smith P.F. Molecular mechanisms of recovery from vestibular damage in
mammals: recent advances // Prog. Neurobiol. 2000
Oct; 62(3): 313-325.
302.
Davidson R.J. Anxiety and affective
style: role of prefrontal cortex and amygdale // Biol Psychiatry. 2002
Jan 1;51(1):68-80.
303. Devi
R.S., Namasivayam A. Modulation of specific immunity by ventral hippocampal
formation in albino rats // J.Neuroimmunol. 1991 Jul; 33(1); 1-6.
304. Devi
R.S., Namasivayam A., Prabhakaran K. Modulation of nonspecific immunity by
hippocampal stimulation // J.Neuroimmunol. 1993 Feb.; 42 (2): 193-197.
305. Divac,
I. Neostriatum and functions of prefrontal cortex. Acta
Neurobiol. Exp. 1972. 33. - P. 461-477.
306. Dowling
G.A., Diamond M.C., Murphy G.M. et al. A morphological study of male rat
cortical asymmetry // Exper. Neurol. 75, 1982. -P51-67.
307. Gainotti
G. Laterality of affect: the emotional behavior of right and left-brain-damaged
patients. In: M.S. Myslobodsky, Editor, Hemisyndromes: Psychopathology,
Neurology, and Psychiatry, Academic Press, New York (1983), pp. 175-192.
308. Glees
P., Cole J., Whitty C.W., Cairns H. The effects of lesions in the cingular
gyrus and adjacent areas in monkeys // J.Neurol.Neurosurg.Psychiatry. 1950 Aug;
13(3): 178-190.
309. Glick
S.D, Cox R. Differential effects of unilateral and bilateral caudate lesion on
side preferences and turning behavior in rats. -J.Compar. and Physiol.Psychol.,
1976, v.90, №5, p.528.
310. Glick
S.D, Jerussi T.P., Zimmerberg B. Behavioral and neuropharmacological correlates
of nigrostriatial asymmetry in rats // Lateralization in the nervous system /
Ed. S.T. Hernad
et al., N.Y.: Acad.press,
1977. p.213-249.
311. Glick
S.D., Weaver L.M., Meibach R.C. Lateralization of reward in rats: differences
in reinforcing thresholds // Science, 1980. V.207. №4435. -
P.1093-1094.
312. Gliddon
C.M., Darlington C.L., Smith P.F. Activation of the
hypothalamic-pituitary-adrenal axis following
vestibular deafferentation in pigmented guinea pig // Brain Res. 2003 Feb 28; 964(2):306-10.
313. Gray
J.A. The psychophysiological nature of introversion-extraversion: A
modification of Eysenck's theory // Biological bases of individual behaviour.
N.Y.: Acad.Press, 1972. - P.182-205
314.
Gulyaeva N., Thompson C., Lazareva
N., Onufriev M., Stepanichev M., Moiseeva Y., Fliss H., Hakim A.M. Tongue
protrusion: a simple test for neurological recovery in rats following focal
cerebral ischemia // J.Neurosci. Methods. 2003. V.125,
№
1-2. - P.183-93.
315.
Haas H.L. Recording from human
hippocampus in vitro // Neurosci. Lett. 1987, Suppl.29.
-
P.13-20.
316. Harley
C.W. A role for norepinephrine in arousal, emotion and learning?: limbic
modulation by norepinephrine and the Kety hypothesis // Prog Neuropsychopharmacol
Biol Psychiatry. 1987;11(4):419-58.
317. Harmon-Jones
E. Early Career Award. Clarifying the emotive functions of asymmetrical frontal
cortical aqctivity // Psycophysiology. 2003. V.40.
N6.- P. 838-848.
318.
Heynen A.J., Bilkey D.K. Induction
of RSA-like oscillations in both the in-vitro and in-vivo hippocampus //
Neuroreport. 1991 Jul;2(7):401-404.
319. Hiroshi
I., Yoshihiko N., Toru M. Neurophisiology of ischemic facial nerve paralysis in
an animal model // Acta oto-laryngol. 1991/ V.111.
№5.-P.
934-942.
320. Jenkins L.W., Moszynski
K., Lyeth B.G., Lewelt W., DeWitt D.S.,
Allen A., Dixon C.E., Povlishock J.T., Majewski
T.J., Clifton G.L. et al. Increased vulnerability of the mildly traumatized rat
brain to cerebral ischemia: the use of controlled secondary ischemia as a
research tool to identify common different mechanisms contributing to
mechanical and ischemic injury. Brain Res. 1989. V. 477. № 1-2. -P.211-224.
. Jorge R., Robinson
R.G. Mood disorders following traumatic brain
injury // Int Rev Psychiatry. 2003
Nov;15(4):317-327.
. Jorge R.E., Robinson
R.G., Moser D., Tateno A., Crespo-Faccoro B., Arndt S. Major depression
following traumatic brain injury //Arch Gen
Psychiatry. 2004 Jan; 61(1):42-50.
323. Kaada
B.R., Pribram K.H., Epstein J.A. Respiratory and vascular responses in monkeys
from temporal pole, insula, orbital surface and cingulate gyrus // J.
Neurophysiol. 1949 V.12. N5.
- P.347.
324. Karczmar
A.G., Richardson D.L., Kindel G. Neuropharmacological and related aspects of
animal aggression. Progr.Neuro-Psychopharmacol. 1978.
V2. №5-6. -P.611-631.
325. Kaufman G.D., Anderson
J.H., Beitz A.J. Brainstem Fos expression following acute unilateral
labyrinthectomy in the rat // Neuroreport. 1992:
3(10): 829-32.
326. Kaufman
G.D., Shindler M.E., Perachio A.A. Correlation of Fos expression and circling
asymmetry during gerbil vestibular compensation // Br.Res. 817 (1999). 246-255.
327. Kim
H.J., Cokeror N.J., Henley C.M. Polyamines increase in the brain stem and
cerebellum following labyrinthectomy. AmJ.Otol. v.18, N2,
1997. - P.214-222.
328. Kim
M.S., Jin B.K., Chun S.W., Lee S.H., Kim J.H., Park B.R. Effect
of C-Fos-like protein expression in the medial
vestibular nucleus at early stage of vestibular compensation in
uvulonodullectomized rats. Neurosci.Lett. 1997 Aug. 15; 231 (3); 147-150.
329. King F.A. Effects of
septal and amygdaloid lesions on emotional behavior and conditioned avoidance
responses in tha rat. J.Nerv. and Mental.
Diseases. 1958, 126; 57-63.
. Kitahara T., Saika T.,
Takeda N., Kiyama H., Kubo Changes in Fos and Jun expression in the rat
brainstem in the process of vestibular compensation. Acta
Otolaryngol.Suppl.Stockl. 1995a. 520Pt 2: 401-404.
331.
Kitahara T., Takeda N., Saika T.,
Kubo T., Kiyama H. Effect of MK801 on Fos expression in the rat brainstem after
unilateral labyrinthectomy. Brain Res. 1995b. Nov 27;
700 (1-2) : 182-190.
332.
Kitahara T., Takeda N., Emson PC,
Kubo T., Kiyama H. Changes in nitric oxide synthese-like immunoreactivities in
unipolar brush cells in the rat cerebellar flocculus after unilateral
labyrinthectomy. Brain Res. 1997a Aug 8; 765 (1) : 1-6.
333. Kitahara
T., Takeda N., Saika T., Kubo T., Kiyama H. Role of the flocculus in the
development of vestibular compensation: immunohistochemical studies with
retrograde tracing and flocculectomy using Fos expression as a marker in the
rat brainstem. Neuroscience. 1997b Jan; 76 (2) : 571- 580.
334. Kolb
B. Animal models for human PFC-related disorders. Review //Prog.Brain Res.
1990; V.85. - P. 501-519
335. Krieg
W.J.S. Accurate placement of minute lesions in the brain of the Albino Rat
Quart // Bull. Of Northwestern Univ. Med Scool. - 1946. V.20.
№2/ - P. 199-208.
336. Kristofikova
Z., Stastny F., Bubenikova V., Druga R., Klaschka J., Spaniel F. Age- and
sex-dependent laterality of rat hippocampal cholinergic system in relation to
animal models of neurodevelopmental and neurodegenerative disorders. Neurochem
Res. 2004
Apr;29(4):671-80.
337. Krutki
P., Jankowska E., Edgley S.A. Are crossed actions of reticulospinal and
vestibulospinal neurons on feline motoneurons mediated by the same or separate
commissural neurons? // J. Neurosci. 2003 sep
3;
23(22): 8041-50.
338. Li Y.X., Tokuyama W.,
Okuno H. et al. Differential induction of brain- derived neurotrophic factor
mRNA in rat inferior olive subregions following unilateral labyrinthectomy //
Neurosci. 2001. V.106. N
2. - P.
385-94.
339.
Lofwing B. Cardiovascular adjiument
induced from rostral cingulated gyrus // Acta Physiol.Scand. 1961. V.63. Suppl.
184.
- P.1-82.
340. McMasters
R.E., Weiss A.H., Carpenter M.B. Vestibular projections to the nuclei of the
extraocular muscles. Degeneration resulting from discrete partial lesions of
the vestibular nuclei in the monkey // Am J Anat. 1966 Jan;118(1):163-94.
341. Maguire
E.A., Frackwiak R.S., Frith C.D. Recalling routes around London activation of
the hippocampus in taxi drivers // J.Neurosci. 1997 Sep 15; 17(18): 7103-7110
342. Maguire
F.A., Frith C.D. Lateral asymmetry in the hippocampal response to the
remoteness of autobiographical memories // J.Neurosci.
2003, 23 (12): 5302-5307
343.
Matsumoto K., Ueda S., Hashimoto T.,
Kuriyama K. Ischemic neuronal injury in the rat hippocampus following forebrain
ischemia: evaluation using in vivo microdialysis // Brain Res. 1991. V.543. N2.
- P.236-242.
344. Matsuyama
K., Drew T. Vestibulospinal and reticulospinal neuronal activity during
locomotion in the intact cat.I.Walking on a level surface // J.Neurophysiol.
2000. 84(5)
- P.2231-2256.
345. Maurice
N., deiau J-M., Glowinski J., Thierry A-M. Relationships between the Prefrontal
Cortex and the Basal Ganglia in the Rat: Physiology of the Cortico-Nigral
Circuits // J.Neuroscience, June 1, 1999, 19(11): 4674-4681.
346. McKenna
J.E., Whishaw I.Q. Complete compensation in skilled reaching success with
associated impairments in limb synergies, after dorsal column lesion in the rat
// J.Neurosci., 1999, 19(5): 1885-1894.
347. Morrison
J.H., Molliver M.E., Grzanna R. Noradrenergic innervation of cerebral cortex:
widespread effects of cortical Lesions. Science, 1979, 205,
N4403. - P.313-316.
348. Moor
R.Y., Bloom F.E. Central Catecholamine Neuron System: Anatomy and Physiology of
the Norepinephrine and Epintphrine Systems. Ann. Rev. Neurosci. N
2. 1979. - C.113- 68.
349. Nauta
W.J.H. Neural associations of the frontal cortex // Acta Neurobiol.Exp., 1972.
V.32, N2.
- P.125-140.
350. Ongur
D., Price J.L. The organization of networks within the orbital and medial
prefrontal cortex of rats, monkeys and humans. Cereb Cortex. 2000
Mar;10(3):206-219.
351.
Pan Q., Long J. Lesions of the
hippocampus enhance or depress humoral immunity in rats // Neuroreport. V.4,
№7, 1993. - P.864-866/
352. Pavlovits
M., Brownstein M.J. Maps and Guide to Microdissection of the Rat Brain. N.Y.,
Amsterdam, London: Elsevier, 1988. 83 p.
353.
Pedraza O., Bowers D., Gilmore R.
Asymmetry of the hippocampus and amygdale in MRI volumetric measurements of
normal adalts // J.Int.Neuropsychol.Soc. 2004. 10 (5):
664-678.
354.
Rath S.A., Klein H.J. Current
applications of the EEG in the copmatose neurosurgical patient // Am.J.EEG
Technol. 1991, 31. - P.65-81.
355. Roberts
W.W., Dember W.N., Brodwick M Alternation and exploration in rats hippocampal
lesions // J.Comp.PhysiolPsychol. 1962, V.55, N 5. -P.695-700.
356. Salzer
T.A., Coker N.J., Henley C.M. Polyamine changes in the vestibular nuclei of
guinea pigs following labyrinthectomy. Am.J.Otol. 1994
Nov; 15(6): 728-34.
357.
Sarkisian V.N. Input-output
relations of Deiters' lateral vestibulospinal neurons with different structures
of the brain // Arch.Ital.Biol. 2000. 138(4).- 259-353.
358. Schoenbaum
G., Chiba A.A., Gallagher M. Neural Encoding in orbitofrontal Cortex and
basolateral amygdale during olfactory discrimination Learning // J.Neurosci.,
1999, 19 (5): 1876-1884.
359. Schuerger
R.J., Balaban C.D. Organization of the coeruleo-vestibular pathway in rats,
rabbits, and monkeys // Brain Res.Brain Res. Rev. 1999.
20(2).
- P.189-217.
360. Schwarting R.C., Huston
J.P. Unilateral 6-hydroxydopamine lesions of meso-striatal dopamine neurons and
their physiological sequelae // Prog. Neurobiol.
1996a Jun; 49(3): 215-266.
361. Schwarting
R.C., Huston J.P. The unilateral 6-hydroxydopamine lesion model in behavioral
brain research. Analysis of functional deficits, recovery and treatments //
Prog Neurobiol. 1996b Oct; 50(2-3):275-331.
362. Scott
J.P. The function of emotions in behavioral systems: theory analysis. In:
Emotion, Theory, Research and Experience. N.-Y: Acad.Press. 1980. - P.35-56.
363.
Showers M.J.C. The cingulat gyrus74
additional motor area and cortical autonomic regulator // J.Comp.Neurol. 1959.
V. 112.
N2.- P.232-301.
364. Smith
W.K. The functional significance of the rostral cingulated gyrus as revealed by
its responses to electrical stimulation // J.Neurophysiol.
1945. V.8. N9.- P.241-255
365. Sperry
R.W. In: Biologycal and biochemical bases of behavior. Madison, 1958.
- p 401.
366. Straka
H., Kunkel A., Dieringer N. Spinal compensation for postural deficits after
hemilabyrinthectomy? Neuroreport. 1993 Sep; 4(9): 10711074.
367. Strauss
E., Kosaka B., Wada J. The neurobiological basis of lateralized cerebral
function. A review. Hum Neurobiol. 1983;2(3):115-27.
368. Sullivan
R.M., Gratton A. Lateralized Effects of Medial Prefrontal Cortex Lesions on
Neuroendocrine and Autonomic Stress Responses in Rats // J. Neuroscience, 1999,
19(7): 2834-2840.
369. Sullivan
R.M., Gratton A. Behavioral effects of excitotoxic lesions of ventral medial
prefrontal cortex in the rat are hemisphere-dependent //
Brain Res. 2002 Feb 8;927(1):69-79.
370. Thompson
R. Behavioral Atlas of the Rat Brain. - New York, 1978.
371. Uno
A., Idoux E., Beraneck M., Vidal P.P., Moor L.E., Wilson V.J., Vibert N. Static
and dynamic membrane properties of lateral vestibular nucleus neurons in guinea
pig brain stem slices // J.Neurophysiol. 2003
Sep; 90 (3): 1689-1703.
372. Uys J.D., Stein D.J.,
Daniels W.M., Harvey B.H. Animal models of
anxiety disorders // Curr.Psychiatry Rep. 2003.
5(4). - P274-81.
373.
Van Groen T., Wyss M.J. Species
differences in hippocampal commissural connections: studies in rat, guinea pig,
rabbit and cat // J.Comp. Neurol. 1988. V.267.
-
P.322-334.
374. Verstynen
T., Tierney R., Urbanski T., Tang A. Neonatal novelty exposure modulates
hippocampal volumetric asymmetry in the rat //
Neuroreport. 2001. 12(14): 3019-3022.
375. Vertes
R.P., Colom L.V., Fortin W.J., Bland B.H. Brainstem sites for the carbachol
elicitation of the hippocampal theta rhythm in the rat // Exp. Brain Res., 1993,
96 (3), p419-429.
376.
Walker S.F. Lateralization of
functions in the vertebrate brain: A review // Brit.J.Psychol. 1980. V.71, N3.
- P.329-367.
377.
Ward A.A. The cingular gyrus: area
24 // J.Neurophysiol. 1948. V.11. N1.- P.13-23.
378. Wetmore
L., Green-Johnson J., Gartner J.G., Sanders V., Nance D.M. The effects of
kainic acid-induced lesions in the lateral septal area on cell-mediated immune
function // Brain Behav.Immun. 1994 Dec; 8(4): 341354.
379.
Wittling W. Psychophysiological
correlates of human brain asymmetry: blood pressure changes during lateralized
presentation of an emotionally laden film. // Neuropsychologia. 1990;
28(5):457-470.
380.
Wolf I., Sutin J. Fiber degeneration
after lateral hypothalamic lesions in the rat // J.Comp.Neurol., 1966. V.127, N
2. - P.137-156.
381. Zilles
K., Wree A. Cortex: Areal and laminar structure. In: The rat nervous system.
Ed.by G.Paxinos. Sydney: Academic Press. 1985. V.2. -
P.375-399.