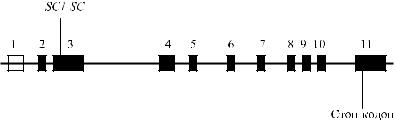
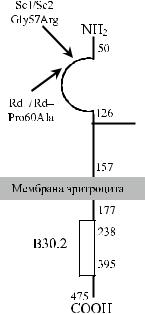
с эритроцитами одного из трех доноров, кровь которых была ему ранее перели-
та (Anderson и соавт. [1]).
Четыре образца анти-Sc2-антител выявлены у доноров – добровольцев, иммунизированных с целью получения реактивов анти-D. Один из доноров, эритроциты которого использовали для иммунизации, имел фенотип D + Sc2 + (Seyfried и соавт. [34]).
По данным Mollison и соавт. [24], из 19 добровольцев с фенотипом D −Sc2 −, получивших по две и более инъекции эритроцитов D + Sc2 +, у 8 образовались анти-D-антитела и только у одного – анти-Sc2-антитела.
Впяти случаях анти-Sc4-антитела вызвали легкую форму ГБН, только одному из новорожденных потребовалось обменное переливание крови
(Rausen и соавт. [32]).
DeMarco и соавт. [6] привели описание одного случая легкой формы ГБН, обусловленной анти-Sc2-антителами.
Вдругом случае у новорожденного была зарегистрирована положительная прямая проба Кумбса, обусловленная антителами анти-Sc1 субкласса IgG3, однако проведениялечебныхмероприятийприэтомнепотребовалось(Kayeисоавт.[14]).
Антитела системы Scianna не вызывали гемолитических осложнений после гемотрансфузий. Однако исследование приживаемости радиоактивно меченных эритроцитов в организме больного, имеющего антитела анти-Sc3, показало относительно быстрое выведение эритроцитов из кровяного русла (McCreary
исоавт. [22]). Интересно, что антитела также вскоре исчезли из плазмы крови реципиента. Попытка стимулировать их последующий синтез путем инъекции эритроцитов Sc:1, −2 эффекта не дала.
Антитела анти-Sc3 у упомянутой выше меланезийской девочки исчезли после спленэктомии и не появлялись даже после трансфузии крови Sc:1, −2 (Woodfield и соавт. [38]). Три образца антител Scianna описали Devine и соавт. [7], об одном из них они сообщили как о причине замедленной гемолитической трансфузионной реакции.
Описано несколько случаев выявления аутоантител анти-Sc1 (Tregellas и со-
авт. [37], McDowell и соавт. [23], Owen и соавт. [28], Pierse и соавт. [30]). У 2
больных, имевших ослабленные антигены Scianna, помимо фиксированных на эритроцитах аутоантител, присутствовали свободно циркулирующие аутоантитела, которые находили в сыворотке крови, но не обнаруживали в плазме
(McDowell и соавт. [23]).
У одного ребенка аутоиммунная гемолитическая анемия, вызванная аутоантителами анти-Sc1, не поддавалась лечению кортикостероидами. Положительный лечебный эффект получен только после удаления селезенки (Owen и соавт. [28]).
Peloquin и соавт. [29] выявили аутоантитела анти-Sc3 в сыворотке крови 2 больных (с лимфомой и болезнью Ходжкина). Экспрессия антигенов Sc1 и Sc3 на их эритроцитах была снижена. Аутоантитела реагировали слабее с эритроци-
тами Sc: −1,2, чем Sc:1, −2.