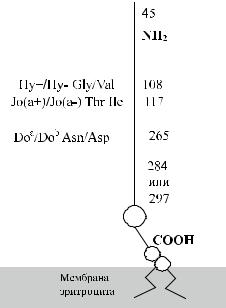
Тесты на приживление эритроцитов in vivo, а также эксперименты с монослоем моноцитов in vitro, показали, что антитела Dombrock ускоряют разруше-
ние эритроцитов, несущих антигены Dombrock (Polesky, Swanson [30], Shirey и соавт. [41]).
Водном случае, описанном Gudino и соавт. [10], у пациента, имевшего анти-Do a-антитела, приживаемость эритроцитов Do(a + ) была нормальной. Переливание ему Do a-положительных эритроцитов не вызвало реакции.
Внекоторых случаях антитела анти-Do a и анти-Do b выявляли post factum, когда трансфузии уже были выполнены. В отдельных случаях эти антитела трактовали как слабые аллогенные, не имеющие клинического значения, ауто-
иммунные или HLA-антитела (Kruskall и соавт. [17], Judd, Steiner [15], Strupp и соавт. [44], Halverson и соавт. [11]).
Впротивоположность антигенам Do a и Do b антиген Gy a более иммуногенен
ивыраженно проявляет себя при разногруппной беременности женщин Gy(a −) (Swanson и соавт. [46], Race, Sanger [32], Clark и соавт. [6], Okubo и соавт. [29]).
Антитела анти-Gy a, анти-Ну и анти-Jo a естественного происхождения не описаны (Daniels [7]). Однако могут быть исключения. Ellisor и соавт. [8] выявили анти-Gy a-антитела у пожилого мужчины, которому не проводили гемотрансфузий. Через 3 мес. антитела исчезли. Эритроциты этого человека, фенотип которого был определен как Gy(a −), адсорбировали анти-Ну-антитела, которые можно было затем элюировать. Стандартные эритроциты Gy(a −) такой способностью не обладали.Этодаетоснованияполагать,чтофенотипGy(a −)можетбытьприобретенным (Reid и соавт. [34]). Процесс подобной трансформации, по-видимому, может сопровождаться появлением соответствующих транзиторных антител.
Антитела анти-Gy a, анти-Ну и анти-Jo a, как правило, относятся к классу IgG (Swanson и соавт. [46], Clark и соавт. [6], Moulds и соавт. [26], Beattie, Castillo [3], Hsu и соавт. [12], Jensen и соавт. [14], Morel и соавт. [24], Brown и соавт. [4], Ellisor и соавт. [8], Barrett и соавт. [2]). Один образец анти-Gy a-антител содержал фракцию IgAи, как удалось установить с помощью антиглобулиновой пробы, связывал комплемент (Clark и соавт. [6]). Один образец анти-Ну-антител, вызывавший прямую агглютинацию эритроцитов, помимо IgG, содержал фрак-
цию IgМ (Barrett и соавт. [2]).
Beattie и Castillo [3] описали случай гемолитической посттрансфузионной реакции у мужчины, имевшего анти-Ну-антитела, которому были перелиты две дозы эритроцитов Ну +.
Описан также реципиент с наличием анти-Gy a-антител, которому были произведены трансфузии 10 доз эритроцитов Gy(a + ) (Mak и соавт. [20]). Какихлибо проявлений несовместимости при этом авторы не наблюдали.
Убольного с транзиторными анти-Gy a-антителами приживаемость эритроцитов Gy(a + ) in vivo была нормальной (Hsu и соавт. [12]). В аналогичных исследованиях ускоренную элиминацию эритроцитов in vivo вызывали антитела анти-Ну (Hsu и соавт. [12]) и анти-Jo a (Viggiano и соавт. [50]).