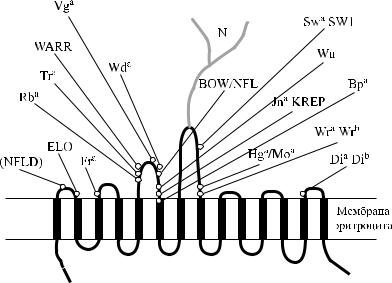
Ассоциация антигенов Diegо с протеином полосы 3 и гликофорином А
Протеин полосы 3 тесно связан в мембране эритроцитов с гликофорином А. Антитела к протеину полосы 3 преципитировали как этот белок, так и гликофорин А (Wainwright и соавт. [169], Tanner [160]). Антитела против гликофорина А существенно снижали подвижность протеина полосы 3 (Nigg и соавт. [125], Che, Sherry [26], Paulitschke и соавт. [130]).
ПомнениюGroves,Tanner[53]гликофоринАспособствуетпереносувновьобразовавшихся молекул белка полосы 3 в мембрану эритроцитов. Однако гликофорин А не является строго обязательной структурой, необходимой для экспрессии белка полосы 3, хотя при его дефиците протеин полосы 3 медленнее движется к мембране и его прикрепление к ней остается незавершенным. В эритроцитах, дефицитных по гликофорину А, протеин полосы 3 имеет более высокую мол. массу и, несмотря на его нормальное количественное содержание, транспорт анионов через мембрану таких эритроцитов затруднен. Эритроциты мышей, у которых направленно был инактивирован ген белка полосы 3 (нокаутные мыши), имели низкое содержание как протеина полосы 3, так и гликофорина А, что дало основание полагать, что белок полосы 3 важен для формирования гликофорина А на мембране эритроцитов (Hassoun и соавт. [57]). Однако эти экспериментальные данные вряд ли можно экстраполировать на человека, поскольку у человека в процессе гемопоэза гликофорин появляется на мембране эритроцитов раньше белка полосы 3 (Southscott и соавт. [156], Daniels, Green [38]). В то же время эритролей-
кемическая линия клеток человека K562 экспрессирует гликофорин А, а протеин полосы 3 не экспрессирует (Gahmberg,Andersson [48], Beckman и соавт. [11]).
Ribeiro и соавт. [142] описали ребенка с полным отсутствием белка полосы 3, однако его эритроциты при этом содержали, хотя и в небольшом количестве, гликофорин А.
Эритроциты трансгенных мышей экспрессировали гликофорин А человека, но в то же время содержание в них мышиного гликофорина А было снижено. Это указывает на возможную конкуренцию двух разных в видовом отношении гликофоринов, участвующих в формировании белка полосы 3 (Auffray и соавт. [10]).
Антиген Wr b не экспрессируется в отсутствие гликофорина А. Эритроциты с дефицитом гликофорина А имеют фенотип En(a −) по системе MN и Wr(a −b −) по системе Diegо (Issitt и соавт. [68, 69]). Образцы эритроцитов Wr(a −b −), которые содержали некоторое количество гибридных гликофоринов, реагировали с антителами анти-En a. Эритроциты индивидов Wr(a +b −) имели нормальную экспрессию антигенов M, N, S, s и En a (Dahr и соавт. [35]). Секвенирование гена GYPA у лицWr(a +b −) не выявило каких-либо особенностей (Bruce и соавт. [17]).
Большинство моноклональных антител к гликофорину А преципитировало указанный гликофорин, но не преципитировало белк полосы 3. Преципитация гликофорина А происходила с анти-Wr b-антителами (Ridgwell и соавт. [144, 145]), которые одновременно преципитировали и протеин полосы 3 (Telen,