Антитела анти-Fy3 впервые обнаруженыAlbrey и соавт. [10] у австралийки Fy(a −b −), имевшей 3 беременности и получавшей гемотрансфузии. Антитела реагировали одинаково интенсивно с эритроцитами Fy(a +b −), Fy(a +b + ) и Fy(a −b + ), не разделялись на анти-Fy a и анти-Fy b дифференциальной адсорбцией. Четыре других найденных образца антител анти-Fy3 реагировали несколько интенсивнее с эритроцитами, содержавшими антиген Fy a (Mannessier
и соавт. [100]).
Антитела анти-Fy3 редко вырабатываются у негроидов, хотя имеются сообщения о выявлении у них нескольких таких образцов (Jensen и соавт. [80], Kosinski и соавт. [85], Molthan, Crawford [114], Oakes и соавт. [125], Oberdorfer
исоавт. [126], Sosler и соавт. [150]). Гораздо чаще у негроидов Fy(a −b −), получавших многократные гемотрансфузии, выявляли антитела анти-Fy a. Тем не менее, антитела Duffy встречаются редко и среди этой категории реципиентов. Так, скрининг сывороток 566 реципиентов-негроидов Fy(a −b −) во Франции ни в одном случае не выявил антител системы Duffy (LePennec и соавт. [89]).
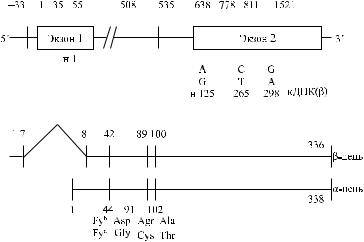
Негроиды Fy(a −b −) гомозиготны по аллелю Fy и мутации в GATA-1 для эритроидных клеток. И хотя их эритроциты имеют фенотип Fy:–3, другие клетки экспрессируют гликопротеины Duffy (Peiper и соавт. [132]). Независимо от мутации в GATA-1 структура генов Fy b и Fy гомологична, поэтому гомозиготы Fy / Fy экспрессируют антигены Fy b и Fy3 в неэритроидных тканях. Это позволяет объяснить редкость антител анти-Fy3, а также анти-Fy b среди негроидов Fy(a −b −). В то же время они способны вырабатывать антитела анти-Fy a. Молекулярная структура фенотипа Fy(a −b −) негроидов, выработавших антитела анти-Fy3, отличается от таковой у негроидов Fy / Fy, не содержащих указанных антител. Более того, антитела анти-Fy3, образовавшиеся у негроидов Fy(a −b −) респондеров, способны улавливать различия в Duffy-гликопротеинах, экспрессированных на разных образцах эритроцитов и неэритроидных клеток.
Антитела анти-Fy3, полученные от негроидов, реагируют слабо или вовсе не реагируют с эритроцитами новорожденных (Kosinski и соавт. [85], Molthan, Crawford [114], Oakes и соавт. [125]), а антитела той же специфичности от лиц других рас реагируют с эритроцитами взрослых и новорожденных одинаково интенсивно (Buchanan и соавт. [23], Mannessier и соавт. [100]). Антигены Duffy
присутствуют на эндотелиальных клетках.
Описаны немедленные и отсроченные гемолитические трансфузионные ре-
акции (Mannessier и соавт. [100], Olteanu и соавт. [129], Vengelen-Tyler [168]), а также случаи легкой ГБН (Kosinski и соавт. [85], Molthan, Crawford [114], Oakes
исоавт. [125]), обусловленные анти-Fy3-антителами. ГБН купировали фототе-
рапией (Albrey и соавт. [10], Buchanan и соавт. [23]).
Мышиные моноклональные антитела анти-Fy3 в серологических реакциях проявляли себя так же, как и поликлональные аллогенные.
Исследования клеток насекомых, подвергнутых трансфекции кДНК, кодирующей гибридные молекулы Fy-гликопротеина и интерлейкина 8, показали, что