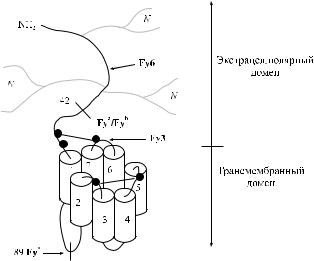
Действие ферментов
Антигены Fy a и Fy b разрушаются большинством протеолитических ферментов: папаином, фицином, бромелином, проназой и химотрипсином. Трипсин не влияет на указанные антигены (Judson, Anstee [83], Miller и соавт. [110], Morton [116]). Обработка эритроцитов сиалидазой также не сказывается на факторах Fy a и Fy b. Ранние сообщения о влиянии трипсина на антигены системы Duffy были обусловлены, как полагают Daniels [43] и Issitt, Anstee [76], искажением результатов из-за примеси химотрипсина.
Хранение нативных эритроцитов в изотоническом растворе хлорида натрия при 12 оС в течение 2 недель приводит к частичой утрате антигенной активности факторов Fy a, Fy b, Fy3 и других Fy-субстанций, специфически ингибирующих соответствующие антитела (Williams и соавт. [179]).
Фенотип Fyx
Фенотип Fy x проявляет себя наличием необычного антигена Fy b, который реагирует не со всеми образцами сывороток анти-Fy b. Специфических антител антиFy x не найдено. Это свидетельствует в пользу того, что антиген Fy x как самостоятельная единица не существует. Тем не менее признак Fy x наследуется кодоми-
нантно, так же как Fy a и Fy b (Chown и соавт. [34, 35], Lewis и соавт. [92]).
Некоторые образцы сывороток анти-Fy a реагируют слабо с эритроцитами Fy aFy x (Chown и соавт. [35], Lewis и соавт. [92]). По адсорбции – элюции высокоактивных анти-Fy b-антител можно отличить лиц, имеющих генотип Fy b / Fy b от лиц, имеющих генотип Fy b / Fy x, однако более четкие результаты получают при использовании ПЦР (Murphy и соавт. [121]).
Фенотип Fy x [Fy(a +b w)] описывали главным образом у лиц белой расы, среди которыхонвстречаетсянестольредко.Среди1108обследованных11человекбыли
Fy(a +b w),1Fy(a −b + w);частотагенаFy x составила0,015(Lewisисоавт.[92]).
Фенотип Fy x описан у индейцев Канады (Buchanan и соавт. [23]). Сообщалось о нескольких лицах, которые были гомозиготны по гену Fy x. Фенотип двоих при первичном тестировании был определен как Fy(a −b −) (Cedergren, Giles [25], Cook и соавт. [39], Habibi и соавт. [58], Lewis и соавт. [92], Parasol и соавт. [131]).
Помимо супрессии антигена Fy b у гомозигот Fy x / Fy x заметно снижена выраженность других часто встречающихся антигенов Duffy: Fy3, Fy5 и Fy6 (Buchanan и соавт. [23], Habibi et al [58, 59], Mallinson и соавт. [99], Marsh [101], Olsson и соавт. [128], Tournamille и соавт. [162], Yazdanbakhsh и соавт. [183]).
Подсчет антигенных участков Fy6 с помощью проточной цитофлюориметрии показал, что их количество на эритроцитах Fy(a −b + ) составляет 2200–2400, на эритроцитах Fy x – от 150 до 250 (Tournamille и соавт. [162]). низкий уровень связывания антител анти-Fy6, обусловленный геном Fy x, наблюдалиTournamille и соавт. [162], Yazdanbakhsh и соавт. [183], используя метод иммуноблоттинга. Авторы полагают, что в присутствии гена Fy x имеет место угнетение синтеза