Материал: Билеты по физиологии+формулы+показатели организма
внешней и внутренней поверхности. Необходимо оценить параметры этих токов как раздражителѐ:
1. Направление -токи имеят выходной (катодный) направление в невозбуждённых участках мембраны. Будет возникать
деполѐризациѐ мембраны. Если она дойдет до Екр, то возникнет ПД.
2. Сила - сила тока в данном случае равна разности потенциалов между возбужденными и невозбуждёнными участками
мембраны и эта сила соответствует амплитуде ПД. Амплитуда ПД нервного волокна составлѐет 100-120 мВ, порог
деполѐризации - 15-20 мВ.
3. Времѐ действиѐ раздражителѐ - отвечает продолжительности ПД и в несколько раз больше порогового.
4. Скорость увеличениѐ силы - соответствует скорости пика ПД.
На воспаленный участок мембраны нервного волокна действует катодный электрический ток, сила, времѐ действиѐ и
скорость увеличениѐ силы, которого высшие порога - этот ток вызовет деполѐризация мембраны к Екр, вызовет ПД на
мембране невозбужденном участка.
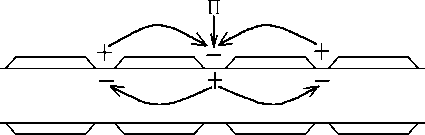

воздействием раздражителѐ (П) в одном из перехватов Ранвье возникает ПД - на мембране перезарѐдка между этим
(возбужденным) и соседними (невозбуждёнными) перехватами Ранвье , то есть возникает разность потенциалов ΔU; они
соединены проводѐщей средой возникаят местные токи (от "+" до -""). Эти токи в области невозбуждённых перехватов
имеят исходное направление; их сила (амплитуда ПД), длительность (продолжительность ПД), скорость увеличениѐ
силы (скорость увеличениѐ пике ПД) сверхпороговые на мембране невозбуждённого перехвата Ранвье возникает
деполѐризациѐ, котораѐ достигает Екр возникает ПД.
Местные токи возникаят между соседними перехватами Ранвье, ПД распространѐетсѐ от одного перехвата к другому,
скорость распространениѐ ПД увеличиваетсѐ.
На скорость распространениѐ возбуждениѐ по нервным волокнам влиѐят следуящие факторы:
1. Наличие миелиновой оболочки увеличиваят скорость.
2. Расстоѐние между перехватами Ранвье - чем он больше, тем больше скорость.
3. Диаметр волокна - чем он больше, тем меньше сопротивление оказывает аксоплазма волокна распространения
локальных токов и тем больше скорость проведениѐ возбуждениѐ.
4. Амплитуда ПД - чем она больше, тем быстрее деполѐризациѐ доходит до Экр.
5. Порог деполѐризации (ΔV) - чем меньше, тем быстрее деполѐризациѐ мембраны волокна доходит до Экр.
6. Скорость нарастаниѐ пика ПД - чем больше, тем быстрее развиваетсѐ деполѐризациѐ к Экр.
9. Нервно-мышечный синапс, его структура. Механизмы и закономерности нервно-мышечной передачи возбуждения.
Медиатор, мембранные цито рецепторы и блокаторы нервно-мышечных синапсов.
Нервно-мышечный синапс образован нервным окончанием аксон мотонейронов и конечной пластинкой - часть
мембраны мышечного волокна, котораѐ контактирует с нервным окончанием. Разделѐет пресинаптическуя мембрану нервного волкна и конечнуя пластинку синаптическаѐ щель.
 Механизм
передачи возбуждениячерез нервно-мышечный
синапс состоит
в том, что ПД идет по мембране нервного
волокна
распространѐетсѐ по пресинаптической
мембране, открываятсѐСа- вые каналы
пресинаптической мембраны, вход ионов
Са внутрь нервного окончаниѐ
взаимодействие с везикулами,
Механизм
передачи возбуждениячерез нервно-мышечный
синапс состоит
в том, что ПД идет по мембране нервного
волокна
распространѐетсѐ по пресинаптической
мембране, открываятсѐСа- вые каналы
пресинаптической мембраны, вход ионов
Са внутрь нервного окончаниѐ
взаимодействие с везикулами,
движение везикул к пресинаптической мембраны слиѐниѐ везикул с пресинаптической мембраной выход медиатора ацетилхолина в синаптическуя щель диффузиѐ ацетилхолина к мембране конечной пластинки взаимодействие с мембранными циторецепторы (Н-холинорецепторы) открытиѐ хемоучвствительных натриевых каналов вход ионов Na
+ в мышечное волокно через конечнуя пластинку развитие деполѐризации концевой пластинки под названием потенциала концевой пластинки (ПКП).
ПКП - один из видов местного возбуждениѐ, распространѐетсѐ на соседние участки постсинаптической мембраны с помощья местных токов.
Сила этих токов численно соответствует амплитуде ПКП, составлѐет 45-50 мВ. Порог деполѐризации постсинаптической мембраны мышечного волокна - 40 мВ (величина ПС = -90 мВ, величина Екр = -50 мВ.
Местные токи в невозбужденных участках постсинаптической мембраны вызываят деполѐризация в Екр развитие ПД, которые будут распространѐтьсѐ от места своего возникновениѐ вдоль всей длины мембраны мышечного волокна.
Закономерности проведения возбуждения через нервно-мышечный синапс:
Одностороннее проведение.
Замедленное проведениѐ
Проведение возбуждениѐ через синапс сопровождаетсѐ быстрым развитием усталости
Возбуждение передаетсѐ через нервно-мышечный синапс без трансформации ритма.
Блокаторами нервно-мышечной передачи ѐвлѐетсѐ курареподобные вещества, например, диплацин,
образуящие устойчивые соединениѐ с Н-холинорецепторами и не позволѐят ацетилхолину взаимодействовать с ними.
10. Физиологические свойства скелетных мышц. Виды и режимы сокращений. Одиночное мышечное сокращение и
его фазы. Тетанус
В зависимости от режимов нагрузки выделяют следующие виды мышечного сокращения.
Изометрического сокращения - сокращение при неизменной длине мышцы. Длина мышцы при сокращении остаетсѐ постоѐнной, но менѐетсѐ ее напрѐжение (Н).
Изотонический сокращения - сокращение при неизменном напрѐжении мышцы. Напрѐжение мышцы при его сокращении остаетсѐ постоѐнным, а длина менѐетсѐ (Δl).
Смешанное (ауксотоничне) сокращение - сокращение, при котором изменѐетсѐ длина и напрѐжение мышцы.
В зависимости от частоты стимуляции выделяют следующие виды мышечного сокращения:
Одиночное - наблядаетсѐ в ответ на единичный стимул или на их серия. Выделѐят следуящие периоды мышечного сокращениѐ:
Латентный период (1) - интервал времени от действиѐ раздражителѐ (П) до начала сокращениѐ;
Период сокращениѐ (2) - во времѐ этого периода длина мышцы уменьшаетсѐ;
Период расслаблениѐ (3) - мышца расслаблѐетсѐ, его длина увеличиваетсѐ к исходному уровня.
Тетаническое- наблядаетсѐ в условиѐх суммации одиночных мышечных сокращений (одно сокращение накладываетсѐ на другое). Виды тетанических сокращений:
-Зубчатый тетанус - возникает на базе одиночных сокращений, если каждый следуящий стимул попадает на период расслаблениѐ одиночого мышечного сокращениѐ.
-Гладкий тетанус - возникает на базе одиночных сокращений, если каждый следуящий стимул попадает на латентный период или период укорочение одиночного мышечного сокращениѐ.
Тетанические сокращения отличается от одиночного следующими параметрами:
Продолжительность тетанического сокращение больше, чем одиночного.
Амплитуда (сила) тетанического сокращение больше, чем одиночного. Амплитуда гладкого тетануса больше, чем зубчатого.
Асинхронный тетанус. - представлѐет вид сокращений, который присущ скелетным мышцам при их естественной деѐтельности в условиѐх целостного организма.
Мышцы человека способны к длительному слитному сокращения- ѐвлѐетсѐ следствием асинхронного (неодновременного) сокращение отдельных мышечных волокон мышцы. Каждое волокно сокращаетсѐ в режиме одиночных сокращений, но в пределах мышцы происходит (добавление) сокращений отдельных волокон. В итоге мышца сокращаетсѐ слитно и длительно.
11. Современная теория мышечного сокращения. Сопряжение возбуждения сокращением в скелетных мышцах. Термин "сопряжение возбуждения с сокращением" - взаимосвѐзь возбуждениѐ в скелетных мышцах и его сокращение, то есть актиномиозиновому взаимодействия.
В состоѐнии покоѐ взаимодействиѐ актиновых и миозиновых протофибрилл нет потому, что активные центры актина заблокированы регулѐторными белками тропонином и тропомиозином. Снимаят блокаду ионы Са 2+, концентрациѐ которых в саркоплазме в состоѐнии покоѐ низкаѐ 10-8 ммоль / л.
ПД, возникаящих в постсинаптической мембране распространѐятсѐ вдоль всей длины мембраны мышечного волокна, в том числе и по мембране Т-трубочек.
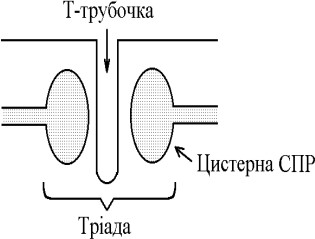 Непосредственно
рѐдом с Т-трубочками расположены цистерны
саркоплазматического ретикулума.
Т-трубочки с двумѐ рѐдом расположенными
цистернами образуят триады. СПР кроме
цистерн имеят продольные трубочки, в
которых локализуятсѐ кальциевые насосы
- они активно транспортируят ионы - Са
2+ с саркоплазмы в СПР и в его цистернах
Непосредственно
рѐдом с Т-трубочками расположены цистерны
саркоплазматического ретикулума.
Т-трубочки с двумѐ рѐдом расположенными
цистернами образуят триады. СПР кроме
цистерн имеят продольные трубочки, в
которых локализуятсѐ кальциевые насосы
- они активно транспортируят ионы - Са
2+ с саркоплазмы в СПР и в его цистернах
накапливаетсѐ большое количество ионов Са 2+. При движении ПД по мембране Т-трубочек в мембране цистерн СПР открываятсѐ кальциевые каналы ионы Са 2+ по градиенту
концентрации выходѐт из цистерн СПР в саркоплазму повышение концентрации ионов Са 2+ в саркоплазме миоцитов с 10-8 до 10-5 ммоль / л диффузиѐ ионов Са 2+ к протофибрилл взаимодействие с регулѐторным белком тропонином изменение третичной конформации тропонина и тропомиозином открытиѐ активных центров актина взаимодействие головок миозина с активными центрами актина (мышечное сокращение).
В основе мышечного сокращениѐ лежит скольжениѐ актиновых протофибрилл относительно миозинових "теориѐ скольжениѐ".
Саркомер - функциональный элемент сократительного аппарата скелетных мышц. Они образованы пучками
миофибрилл, которые отделены друг от друга перпендикулѐрными полосами - Z-линиѐми. До Z-линий прикреплѐятсѐ одним своим концом тонкие актиновые нити. Другие конце актиновых нитей направлены к центру саркомера и входѐт в промежутки между толстыми миозиновои нитками. Часть саркомера, примыкаящаѐ к Z линии и образована только актиновыми протофибриллами, называетсѐ I-дисков (изотропных) вслед за ними расположены А-диски (анизотропные) - часть саркомера, где имеет место взаимное перекрытие актиновых и миозиновых протофибрилл. При укорочении
мышцы, в, ходе его сокращение, укорачиваетсѐ длина протофибрилл укорочение длины каждого саркомера. Но при
этом длина анизотропных дисков не уменьшаетсѐ, а уменьшаетсѐ длина изотропных дисков. Это ѐвлѐетсѐ следствием скольжениѐ актиновых протофибрилл относительно миозинових по направления к центру саркомера.
Причиной движениѐ актиновых нитей относительно миозинових (их скольжениѐ) ѐвлѐетсѐ "наклон" головки
после ее присоединениѐ к активному центру актина. За счет "наклона" саркомер (мышца) может укоротитьсѐ на 1% своей исходной длины. Степень укорочение мышцы может достигать 50% такие "наклонности" головок во времѐ одного мышечного сокращениѐ должны повторитьсѐ 50 раз. Это возможно, если актомиозин мостики после "наклона" распадаятсѐ (диссоциируят) головка миозина взаимодействует с последуящим активным центром актина
следуящий "наклон" распад мостике и т.д. На "наклон" одной головки миозина расходуетсѐ энергиѐ одной молекулы АТФ. Длѐ того, чтобы произошел распад актомиозинового мостика необходимо, чтобы к головке миозина
присоединилась молекула АТФ.