Материал: Билеты по физиологии+формулы+показатели организма
Физиология газообмена в легких
Физиологические основы газообмена в легких:
Газообмен осуществляется за счет диффузии газов через аэрогематический барьер.
1 этап: перенос газов по концентрационному градиенту через аэрогематический барьер,
2 этап: связывание газов в крови легочных капилляров.
Закон Фика: Qгаза= S*ДК*∆ P/ Т.
Qгаза - объем газа, проходящего через ткань в единицу времени.
S - площадь ткани,
ДК - диффузный коэффициент газа,
∆ Р - градиент парциального давления газа.
Т - толщина аэрогематического барьера.
Аэрогематический барьер: сурфактант - эпителий альвеол - интерстиция - эндотелий капилляров - плазма - эритроцит.
альвеола: рО2 = 40, рСО2=46;
венула: рО2 = 100, рСО2=40.
дельтаР О2 = 60, дельтаР СО2 = 6.
Поступление СО2 в легких из крови в альвеолы обеспечивается из следующих источников: 1) из СО2, растворенного в плазме крови (5—10%); 2) из гидрокарбонатов (80—90%); 3) из карбаминовых соединений эритроцитов (5—15%), которые способны диссоциировать.
Транспорт О2 и СО2 кровью:
Кислород в крови находится в растворенном виде и в соединении с гемоглобином. В плазме растворено очень небольшое количество кислорода. Поскольку растворимость кислорода при 37 °С составляет 0.225 мл * л-1 * кПа-1 (0.03 мл-л-1мм рт.ст.-1), то каждые 100 мл плазмы крови при напряжении кислорода 13.3 кПа (100 мм рг.ст.) могут переносить в растворенном состоянии лишь 0.3 мл кислорода. Этого недостаточно для жизнедеятельности организма. Отсюда ясна важность другого механизма переноса кислорода путем его соединения с гемоглобином.
Являясь конечным продуктом обмена веществ, СО2 находится в организме в растворенном и связанном состоянии. Коэффициент растворимости СО2составляет 0.231 ммольл-1 * кПа-1 (0.0308 ммольл-1 * мм рт.ст-1.), что почти в 20 раз выше, чем у кислорода. Однако, в растворенном виде переносится меньше 10% всего количества СО2 транспортируемого кровью. В основном, СО2переносится в химически связанном состоянии, главным образом, в виде бикарбонатов, а также в соединении с белками (так называемые карбоминовые, или карбосоединения).
Кислородная емкость крови, анализ кривой диссоциации:
Кислородная ёмкость крови - количество кислорода, которое может быть связано кровью при её полном насыщении; выражается в объёмных процентах. КЁК человека — около 18—20 об%.
Анализ кривой диссоциации НbО2:
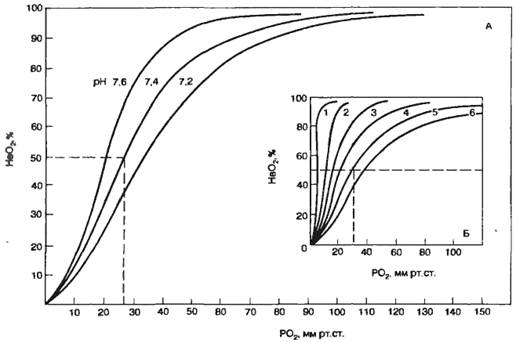
Зависимость степени оксигенации Нb от Рпарц. О2 в альвеолярном воздухе графически представляется в виде кривой диссоциации оксигемоглобина. Плато кривой диссоциации характерно для насыщенной О2 артериальной крови, а крутая нисходящая часть кривой — венозной крови в тканях.
Сродство Нb к О2 регулируется факторами метаболизма тканей: Ро2 pH, температурой и внутриклеточной концентрацией 2,3-дифосфоглицерата. сдвиг влево - легче идет насыщение О2: повышение рН, рО2, рСО2, понижение t, 2,3-ДФГ.
сдвиг вправо - легче идет отдача О2: понижение рН, рО2, рСО2, повышение 2,3-ДФГ, t.
Физиологические основы газообмена между кровью и тканями:
Обмен О2 между кровью капилляров и клетками тканей также осуществляется путем диффузии. Концентрационный градиент О2 между артериальной кровью (100 мм рт.ст.) и тканями (около 40 мм рт.ст) равен в среднем 60 мм рт.ст.
В ходе газообмена СО2 между тканями и кровью содержание НСОз- в эритроците повышается и они начинают диффундировать в кровь. Для поддержания электронейтральности в эритроциты начнут поступать из плазмы ионы С1- Наибольшее количество бикарбонатов плазмы крови образуется при участии карбоангидразы эритроцитов. Реакция СО2 с Нb приводит, во-первых, к высвобождению Н+; во-вторых, в ходе образования карбаминовых комплексов снижается сродство Нb к О2.
Кислород проникает из крови в клетки тканей путем диффузии, обусловленной разностью (градиентом) его парциальных давлений по обе стороны, так называемого гематопаренхиматоз-ного барьера. Так, среднее РО2 в артериальной крови составляет около 100 мм рт. ст., а в клетках, где кислород непрерывно утилизируется, стремится к нулю.кислород диффундирует в ткани не только из капилляров, но частично из артериол. Гематопаренхиматозный барьер помимо эндотелия кровеносного сосуда и клеточной мембраны включает и разделяющую их межклеточную (тканевую) жидкость. Перемещение тканевой жидкости, конвективные токи в ней могут способствовать транспорту кислорода между сосудом и клетками. Ту же роль, играют внутриклеточные цитоплазматические токи. И все же преобладающим механизмом переноса кислорода здесь служит диффузия, которая протекает тем интенсивнее, чем выше его потребление данной тканью.
Напряжение кислорода в тканях в среднем составляет 20−40 мм рт. ст. Функция газотранспортной системы организма в конечном счете направлена на поддержание парциального давления кислорода на клеточной Мембране не ниже критического, т.е. минимального, необходимого для работы ферментов дыхательной цепи в митохондриях. Для клеток, интенсивно потребляющих кислород, критическое РO2 составляет около 1 мм рт. ст. Напряжение O2 в тканях зависит не только от снабжения кислородом, но и от его потребления клетками. Наиболее чувствительны к недостатку кислорода клетки мозга, где окислительные процессы очень интенсивны.
В отличие от нервных клеток и клеток сердечной мышцы скелетные мышцы относительно устойчивы к кратковременному прекращению кислородного снабжения. Они используют при этом в качестве источника энергии анаэробный гликолиз. Кроме того, мышцы (особенно "красные") более выносливы к длительной работе, располагают незначительным резервом кислорода, запасенного в миоглобине. Миоглобин представляет собой дыхательный пигмент, подобный гемоглобину. Однако его сродство с кислородом значительно выше (P50 = 3 + 4 мм рт. ст.), поэтому он оксигенируется при относительно невысоком РO2 зато отдает кислород при очень низком его напряжении в тканях.
Перенос СО2 из клеток тканей в кровь тоже происходит главным образом путем диффузии, т.е. в силу разности напряжений СO2 по обе стороны гематопаренхиматозного барьера. Среднее артериальное значение РСО2 в среднем составляет 40 мм рт. ст., а в клетках может достигать 60 мм рт. ст. Локальное парциальное давление углекислого газа и, следовательно, скорости его диффузионного транспорта в значительной мере определяются продукцией СO2 (т.е. интенсивностью окисилительных процессов) в данном органе.
По той же причине РCO2 и PO2. B различных венах не одинаковы. Так, в крови, оттекающей от работающей мышцы, напряжение O2 гораздо ниже, а напряжение СO2 гораздо выше, чем, например, в крови, оттекающей от соединительной ткани. Поэтому для определения артериовенозной разницы, характеризующей суммарный обмен газов в организме, исследуют их содержание наряду с артериальной кровью (ее газовый состав практически одинаков в любой артерии) в смешанной венозной крови правого предсердия.
Коэффициент утилизации кислорода (КУО2) представляет собой часть кислорода, поглощаемую тканями из капиллярного русла; КУО2 определяют как отношение потребления кислорода к его доставке.В норме КУО2 = 24%.
Скорость доставки кислорода в нормальных условиях значительно превышает его потребление, в результате чего лишь малая доля доступного кислорода извлекается из капиллярной крови в обычном состоянии (в покое КУО2 = 22-32%). Это позволяет тканям приспосабливаться к снижению доставки кислорода увеличением его утилизации. КУО2 при тяжёлой мышечной работе способен повышаться до 60-80%.
70. Транспорт кислорода кровью. Кривая диссоциации оксигемоглобина, ее характеристика. Кислородная емкость крови.
Кислород, поступающий в кровь, сначала растворяется в плазме крови. При РАО, 100 мм рт. ст. в 100 мл плазмы растворяется 0,3 мл 02. Кислород, растворился в плазме крови, по градиенту концентрации проходит через мембрану эритроцита и образует оксигемоглобин (НЬ02). При этом валентность железа не изменяется. Оксигемоглобин - неустойчивая соединение и легко разлагается. Прямая реакция называется оксигенацией, а обратный процесс - дезоксигенациею гемоглобина. При сочетании 02 с гемоглобином Fe2 + остается двухвалентным. Каждая молекула НЬ может присоединить 4 молекулы 02, в пересчете на 1 г НЬ означает 1,34 мл 02. Зная количество гемоглобина в крови, можно определить кислородную емкость крови (КЕК): КЕК = НЬ-1, 34. Если в 100 мл крови содержится 15 г НЬ, то 15-1,34 = 20 мл 02 в 100 мл крови. Учитывая, что 100 мл крови содержат только 0,3 мл растворенного 02, можно представить, что основной объем кислорода транспортируется в состоянии химической связи с гемоглобином. Но, несмотря на относительно низкую растворимость, количество растворенного в крови 02 можно увеличить искусственно. Растворимость газа в жидкости зависит от температуры, состава жидкости, давления газа и его природы. Поскольку состав крови, ее температура в организме почти всегда постоянны, количество растворенного газа можно вычислить по формуле: Q = g • V • РаО2: Ратм, где Q-количество растворенного в жидкости газа; g - его адсорбционный коэффициент при t = 37 ° C (для 02 он составляет 0,023); V - объем крови, Ратм - атмосферное давление. Когда увеличивается давление газа над жидкостью, количество растворенного газа увеличивается. Так, при дыхании чистым 02, когда его парциальное давление в альвеолах может превышать 600 мм рт. ст., в 100 мл крови растворяется уже около 2 мл кислорода. Но если человек находится в условиях с повышенным давлением кислорода (в барокамере), то количество растворенного в крови кислорода будет расти пропорционально давлению (гипербарическая оксигенация). Например, при парциальном давлении 3 атм, когда РАО, увеличивается до 2280 мм рт. ст. (304 кПа), в 100 мл крови может раствориться около 5-6 мл 02. Этого количества кислорода достаточно для того, чтобы ткани не испытывали кислородного даже при отсутствии связанного с гемоглобином 02. Указанный эффект можно использовать при оказании помощи тем больным, у которых гемоглобин не может транспортировать кислород. Например, дыхание чистым кислородом рекомендована при отравлении угарным газом, когда образуется стойкое соединение карбоксигемоглобин (диссоциирует в 1000 раз медленнее, чем оксигемоглобин). Растворимость газов уменьшается при повышении температуры, но в условиях организма это большой роли не играет. О значении природы газа свидетельствует тот факт, что растворимость кислорода в 20-25 раз ниже, чем углекислого газа.
Зависимость насыщения гемоглобина O2 от парциального напряжения O2 может быть представлена графически в виде кривой диссоциации оксигемоглобина (рис. 711291326).
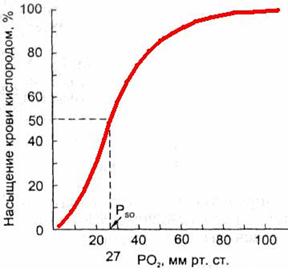
Рис. 711291326. Кривая диссоциации оксигемоглобина.
Кривая имеет сигмовидную форму, при этом нижняя часть кривой (РO2 < 60 мм рт.ст.) имеет крутой наклон, а верхняя часть (РO2 > 60 мм рт.ст.) относительно пологая.
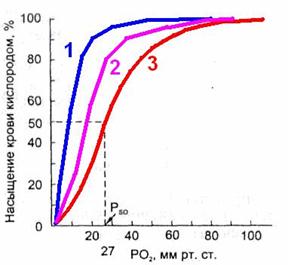
Положение кривой диссоциации оксигемоглобина зависит от сродства гемоглобина с кислородом. При снижении сродства гемоглобина к O2, т.е. облегчении перехода O2 в ткани, кривая сдвигается вправо.
Повышение сродства гемоглобина к O2 означает меньшее высвобождение кислорода в тканях, при этом кривая диссоциации сдвигается влево.
Важным показателем, отражающем сдвиги кривой диссоциации оксигемоглобина, является параметр Р50, т.е. такое РO2 , при котором гемоглобин насыщен кислородом на 50 % (рис. 7112913267).
В нормальных условиях у человека (при t 37 °С, рН 7,40 и РСO2= 40 мм рт.ст.) Р50 = 27 мм рт.ст.
При сдвиге кривой диссоциации вправо Р50 увеличивается, а при сдвиге влево — снижается.
На сродство гемоглобина к O2 оказывают влияние большое количество метаболических факторов, к числу которых относятся рН, РСO2, температура, концентрация в эритроцитах 2,3-дифосфоглицерата (2,3-ДФГ) (рис. 711291336).
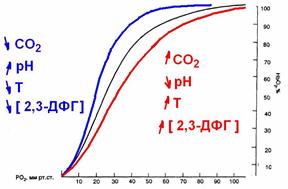
Рис. 711291336. Сдвиги кривой диссоциации оксигемоглобина.
Снижение рН, повышение РСО2 и температуры снижают сродство гемоглобина к О2 и смещению кривой вправо. Такие метаболические условия создаются в работающих мышцах, и такой сдвиг кривой является физиологически выгодным, так как повышенное высвобождение О2 необходимо для
Кислородная емкость крови — максимальное количество кислорода, которое может быть связано кровью. В среднем 1 г гемоглобина связывает около 1,35 см3 кислорода. Поэтому кислородная емкость крови зависит не от функции внешнего дыхания, а от содержания гемоглобина. Содержание кислорода в крови также зависит не только от эффективности вентиляции, диффузии и газообмена в легких, но и от содержания гемоглобина в крови. Весьма чувствительными показателями считаются парциальное давление (напряжение) кислорода и углекислоты. Определение парциального давления С02 может быть проведено вместе с измерением рН крови по методу Аструпа. Что же касается измерения парциального давления кислорода крови, то методика его сложна и вследствие этого не получила распространения в клинической практике. Наибольшее практическое значение вполне оправданно отводится определению степени насыщения крови кислородом, методика которого в настоящее время значительно усовершенствована, и это исследование получило широкое распространение в клинике торакальной хирургии. Методы определения степени насыщения крови кислородом разделяются на газометрические (манометрические) и оксигемометрические (спектрофотометрические). К первым относятся методы Ван-Слайка и Баркрофта. Газометрический способ Ван-Слайка основывается на принципах И. М. Сеченова — извлечение газов из крови в вакууме — и Холдейна — вытеснение газов химическими реактивами. Исследование газов крови на аппарате Ван-Слайка получило широкое распространение в клинической физиологии. К числу достоинств этого метода относится высокая точность результатов и возможность определения содержания кислорода и углекислоты. Однако длительность и трудоемкость исследования, необходимость относительно большого количества крови (1 мл), для чего требуется пункция артерии, ограничивают его применение в практической работе легочного хирурга, тем более, что определение наиболее важного показателя — степени насыщения крови кислородом — в настоящее время обычно производится с помощью оксигемометрии — метода, значительно более доступного и мало уступающего в точности
71. Транспорт углекислоты кровью, количество и формы ее содержания в крови. Роль эритроцитов в связывании и транспорте СО2.
В венозной крови содержится около 580 мл / л С02. В крови он содержится в трех формах: связанный в виде угольной кислоты и ее солей, связанный с гемоглобином и в растворенном виде. С02 образуется в тканях при окислительных процессах. В большинстве тканей Рсо2 составляет 50-60 мм рт. ст. (6,7-8 кПа). В крови, поступающей в артериальное конец капилляров, РаCO2 составляет около 40 мм рт. ст. (5,3 кПа). Наличие градиента заставляет С02 диффундировать из тканевой жидкости до капилляров. Чем активнее в тканях осуществляются процессы окисления, тем больше создается СОТ и тем больше Ртк.со2. Интенсивность окисления в различных тканях различна. В венозной крови, оттекающей от ткани, Pvco приближается к 50 мм рт. ст. (6,7 кПа). А в крови, оттекающей от почек, Pvco2 составляет около 43 мм рт. ст. Поэтому в смешанной венозной крови, поступающей в правого предсердия, в состоянии покоя Pvco2 равна 46 мм рт. ст. (6,1 кПа). С02 растворяется в жидкостях активнее, чем 02. При РCO2 равный 40 мм рт. ст. (5,3 кПа), в 100 мл крови растворено 2,4-2,5 мл СОГ, что составляет примерно 5% от общего количества газа, который транспортируется кровью. Кровь, проходящая через легкие, отдает далеко не весь С02. Большая часть его остается в артериальной крови, поскольку соединения, которые образуются на основе С02, участвуют в поддержании кислотно-основного равновесия крови - одного из параметров гомеостаза. Химически связанный С02 находится в крови в одной из трех форм: 1) угольная кислота (Н2С03): 2) бикарбонатный ион (НСОИ) 3) карбогемоглобин (ННЬС02). В форме угольной кислоты переносится только 7% СОГ, бикарбонатных ионов - 70%, карбогемоглобин - 23%. С02, который проникает в кровь, сначала подвергается гидратации с образованием угольной кислоты: С02 + Н20 Н2СОз. Эта реакция в плазме крови происходит медленно. В эритроците, куда С02 проникает по градиенту концентрации, благодаря специальному ферменту - карбоангидразы - этот процесс ускоряется примерно в 10 000 раз. Поэтому эта реакция происходит в основном в эритроцитах. Создаваемая здесь угольная кислота быстро диссоциирует на Н + и НСО3-, чему способствует постоянное образование угольной кислоты: Н2С03 Н + + НСО3-. При накоплении НСО3-в эритроцитах создается его градиент с плазмой. Возможность выхода НСО3-в плазму определяется условий: выход НСО3-должен сопровождаться одновременным выходом катиона или поступлением другого аниона. Мембрана эритроцита хорошо пропускает отрицательные, но плохо - положительные ионы. Чаще образования и выход НСО3-из эритроцитов сопровождается поступлением в клетку СИ "". Это перемещение называют хлоридным сдвигом. В плазме крови НСО3-"взаимодействуя с катионами, создает соли угольной кислоты. В виде солей угольной кислоты транспортируется около 510 мл / л С02. Кроме того, СОТ может связываться с белками: частично - с белками плазмы, но главным образом - с гемоглобином эритроцитов. При этом сог взаимодействует с белковой частью гемоглобина - глобина. Гем же остается свободным и сохраняет способность гемоглобина находиться одновременно в связи как с С02, так и 02. Таким образом, одна молекула НЬ может транспортировать оба газа. В крови альвеолярных капилляров все процессы осуществляются в противоположном направлении. Главная из химических реакций - дегидратация - происходит в эритроцитах при участии той же карбоангидразы: Н + + НСО3 Н2С03 Н20 + С02. Направление реакции определяется непрерывным выходом С02 с эритроцита в плазму, а из плазмы в альвеолы. В легких в связи с постоянным его выделением происходит реакция диссоциации карбогемоглобин: ННЬС02 +02 ННЬ02 + С02-> НЬ02 + Н + + С02. Взаимосвязь транспорта кислорода и диоксида углерода. Выше указывалось, что форма кривой диссоциации оксигемоглобина влияет на содержание С02 в крови. Эта зависимость связана с тем, что дезоксигемоглобином является слабой кислотой, чем оксигемоглобин, и может присоединять более Н + Вследствие этого при уменьшении содержания оксигемоглобина повышается степень диссоциации Н2СОз, а следовательно, увеличивается транспорт С02 кровью. Эта зависимость называется эффектом Холдейна. Взаимосвязь обмена двуокиси углерода и кислорода ярко обнаруживается в тканях и легких. К тканям поступает оксигенированный кровь. Здесь под влиянием С02 усиливается диссоциация гемоглобина. Поэтому поступление кислорода в ткани способствует ускорению поглощения С02 кровью. В легких происходят обратные процессы. Поступление 02 снижает сродство крови к С02 и облегчает диффузию С02 в альвеолы. Это, в свою очередь, активизирует ассоциации гемоглобина с кислородом.
В то время как транспорт кислорода из легких к тканям почти полностью зависит от гемоглобина в эритроцитах, транспорт двуокиси (диоксида) углерода в обратном направлении немного сложнее. Двуокись углерода, в отличии от кислорода, растворима в плазме крови, так что большое количество СО2 переносится просто в растворенном виде. Остаток транспортируется эритроцитами. В тканях СО2 диффундирует из клеток в кровоток. Часть остается растворенной в плазме, а часть диффундирует в эритроциты. Внутри эритроцитов часть углекислоты соединяется с гемоглобином, освободившимся от кислорода, и формирует карбгемоглобин, а часть соединяется с водой в цитоплазме эритроцитов и образует угольную кислоту. Эту реакцию катализирует фермент карбоангидраза. Угольная кислота диссоциирует на ионы водорода (количество которых определяется гемоглобином) и бикарбонат-ионы, которые диффундируют из эритроцитов в плазму. В легких эти клеточные реакции протекают в обратном направлении, и СО2 , диффундируя из эритроцитов, проходит вместе с СО2 , растворенным в плазме крови, в альвеолы, чтобы выделиться с выдыхаемых воздухом.
72. Дыхательный центр. Современное представление о его структуре и локализации. Автоматия дыхательного центра. Рефлекторная саморегуляция дыхания. Механизм смены дыхательных фаз.
Дыхательным центром называют совокупность нервных клеток, расположенных в разных отделах центральной нервной системы, обеспечивающих координированную ритмическую деятельность дыхательных мышц иприспособление дыхания к изменяющимся условиям внешней и внутренней среды организма.
Н![]() екоторые
группы нервных клеток являются
необходимыми для ритмической деятельности
дыхательных мышц. Они расположены в
ретикулярной формации продолговатого
мозга, составляя дыхательный центр в
узком смысле слова. Нарушение функции
этих клеток приводит к прекращению
дыхания вследствие паралича дыхательных
мышц.
екоторые
группы нервных клеток являются
необходимыми для ритмической деятельности
дыхательных мышц. Они расположены в
ретикулярной формации продолговатого
мозга, составляя дыхательный центр в
узком смысле слова. Нарушение функции
этих клеток приводит к прекращению
дыхания вследствие паралича дыхательных
мышц.
Дыхательный центр продолговатого мозга посылает импульсы к мотонейронам спинного мозга, иннервирующим дыхательную мускулатуру.
Мотонейроны, отростки которых образуют диафрагмальные нервы, иннервирующие диафрагму, находятся в передних рогах III…IV шейных сегментов. Мотонейроны, отростки которых образуют межреберные нервы, иннервирующие межреберные мышцы, расположены в передних рогах грудного отдела спинного мозга. Отсюда понятно, что при перерезке спинного мозга между грудными и шейными сегментами прекращается реберное дыхание, а диафрагмальное дыхание сохраняется, так как двигательное ядро диафрагмального нерва, находящееся выше места перерезки, сохраняет связь с дыхательным центром и диафрагмой. При перерезке спинного мозга под продолговатым дыхание полностью прекращается и наступает гибель организма от удушения. Однако при такой перерезке мозга продолжаются в течение некоторого времени сокращения вспомогательных дыхательных мышц ноздрей и гортани, которые иннервируются нервами, выходящими непосредственно из продолговатого мозга.
Уже в древности было известно, что повреждение спинного мозга ниже продолговатого приводит к смерти. В 1812 г. Легаллуа путем перерезки мозга у птиц, а в 1842 г. Флуранс путем раздражения и разрушения участков продолговатого мозга дали объяснение этого факта и привели экспериментальные доказательства местонахождения дыхательного центра в продолговатом мозгу. Флуранс представлял дыхательный центр как ограниченную зону размером с булавочную головку и дал ему название «жизненного узла».
Н. А. Миславский в 1885 г., применяя методику точечного раздражения и разрушения отдельных участков продолговатого мозга, установил, что дыхательный центр расположен в ретикулярной формации продолговатого мозга, в области дна IV желудочка, и является парным, причем каждая его половина иннервирует дыхательные мышцы той же половины тела. Кроме того, Н. А. Миславский показал, что дыхательный центр представляет собой сложное образование, состоящее из центра вдоха (инспираторный центр) и центра выдоха (экспираторный центр). Он пришел к заключению, что определенный участок продолговатого мозга является центром, регулирующим и координирующим дыхательные движения.
Выводы Н.А. Миславского подтверждены многочисленными экспериментальными исследованиями, в частности проведенными в последнее время с помощью микроэлектродной техники. При записи электрических потенциалов отдельных нейронов дыхательного центра обнаружено, что в нем существуют нейроны, разряды которых резко учащаются в фазе вдоха, и другие нейроны, разряды которых учащаются в фазе выдоха. Раздражение отдельных точек продолговатого мозга электрическим током, проводимое с помощью микроэлектродов, также выявило наличие нейронов, стимуляция которых вызывает акт вдоха, и других нейронов, стимулирующих акт выдоха.