|
|
|
|
Порушення функції щитоподібної залози |
дження чи в ранньому дитинстві, відомий як кре- |
|
|
|
|
|
|
Гіпертиреоз |
|
|
|
тинізм. При гіпофункції щитоподібної залози в |
|
|
|
|
|
|
|
|
|
дорослих розвивається мікседема. |
|
|
|
Гіпертиреоз може виникнути при: |
|
|
|
Лікування гіпотиреозу полягає в призначенні |
|
|
|
— надмірній стимуляції залози, як при хворобі |
пероральної замісної терапії тиреоїдним гормо- |
|
Грейвса (базедова хвороба, тиреотоксикоз); |
ном, при первинному гіпотиреозі (найчастіша |
|
|
|
— автономній секреції тироксину, як при ток- |
причина) — нормалізації вмісту тирозинстиму- |
|
сичній аденомі та токсичному багатовузловому |
лювального гормону. |
|
зобі; |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
— надлишковому прийомі екзогенного гормо- |
|
|
ну. |
|
|
|
|
|
|
|
|
|
|
|
|
|
Гормональне регулювання метаболізму |
|
|
|
При надмірному вмісті тиреоїдного гормону |
кальцію та фосфору |
|
зростає потреба клітин у кисні, збільшується про- |
|
|
|
дукція тепла і використання метаболітів, що зви- |
Постійний рівень Са в крові дуже важливий |
|
чайно призводить до зниження маси тіла. Озна- |
для м’язової діяльності, збудливості нервової тка- |
|
ками тиреотоксикозу є схуднення, підвищення |
нини, для скоротливої здатності міокарда, при |
|
температури тіла, інтоксикація недоокисненими |
згортанні крові. |
|
продуктами обміну, ураження серцевої та нерво- |
У регулюванні метаболізму кальцію беруть |
|
вої систем. |
|
|
|
|
|
|
|
|
|
|
|
|
|
участь три гормони: паратгормон, кальцитонін, |
|
|
|
Надлишкова секреція тирозинстимулюваль- |
1,25-дигідроксихолекальциферол. Четвертий гор- |
|
ного гормону (ТСГ) дуже рідко стає причиною |
мон — пептид, подібний до паратгормону (PTHrP) |
|
тиреотоксикозу. |
|
|
|
|
|
|
|
|
|
|
|
|
|
(від англ. pаrаthyrеoid hormones — related pro- |
|
|
|
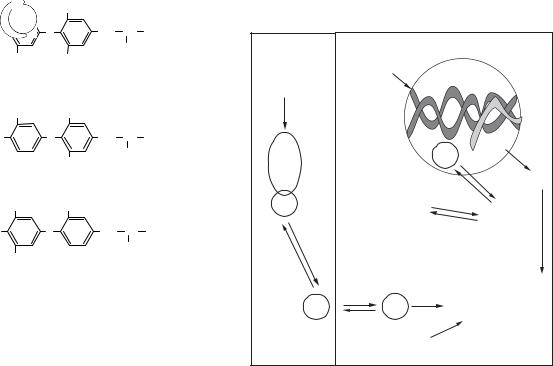
Лікування гіпертиреозу (гіпертироксинемії) |
stein), він діє на один із рецепторів паратгормо- |
|
залежить від його причини і полягає у призна- |
ну і є важливим для розвитку скелетних м’язів in |
|
ченні антитиреоїдних препаратів (метимазолу, |
utero. На метаболізм кальцію також можуть |
|
пропілтіоурацилу), β -блокаторів (пригнічують |
впливати гормони, що регулюють вміст фосфору, |
|
перетворення Т4 на активну форму Т3), радіоак- |
глюкокортикоїди, гормон росту, естрогени та |
|
тивного йоду, проведенні хірургічної операції |
різноманітні фактори росту. |
|
(рис. 15.4). |
|
|
|
|
|
|
|
|
|
|
|
|
|
Кальцій у кістках буває двох типів: легкооб- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
мінний пул і набагато більший пул стабільного |
I |
|
|
|
|
|
|
|
|
T4 |
|
|
T3 |
|
|
|
Метаболічні |
кальцію, здатний лише до повільного обміну. Є |
|
|
|
|
|
|
|
|
|
|
|
ефекти |
дві різні незалежні гомеостатичні системи, що |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
взаємодіють і впливають на кальцій у кістках. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Одна система регулює Са2+ у плазмі, за її до- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Симптоми |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
помогою щодня 500 ммоль Са2+ потрапляє в |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
кістковий пул кальцію, здатний до легкого об- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
міну. |
|
|
Пропілтіоурацил, |
|
|
|
β -блокатори блокують ак- |
|
|
метимазол блокують |
|
|
|
тивацію тиреоїдного гормо- |
|
Друга система пов’язана з ремоделюванням |
|
|
вбудовування йоду |
|
|
|
ну і зменшують симптоми |
|
кістки шляхом постійної взаємодії між резорбцією |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
та утворенням кістки, у дорослих вона на 95 % |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
відповідає за утворення кістки. Проте взаємо- |
|
|
|
|
|
Йодид літію блокує |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
обмін Са2+ між плазмою та цим стабільним пулом |
|
|
|
|
|
вивільнення гормонів |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
становить 7,5 ммоль за добу. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Рис. 15.4. Лікування гіпертиреозу |
Значну кількість кальцію фільтрують нирки, |
|
|
|
|
однак 98–99 % його реабсорбується (близько |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
60 % у проксимальних канальцях, решта — у |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
висхідній частині петлі Генле та дистальних ка- |
|
|
|
|
|
|
Гіпотиреоз |
|
|
|
нальцях нефрону). Реабсорбцію в дистальних |
|
|
|
Гіпотиреоз може виникнути при: |
|
|
|
канальцях регулює паратгормон. |
|
|
|
|
|
|
Паратгормон (паратиреоїдний гормон, полі- |
|
|
|
— недостатності щитоподібної залози (пер- |
|
|
|
пептид) — його секретують паращитоподібні за- |
|
винний гіпотиреоз, |
|
чи автоімунний тиреоїдит |
|
Хашимото); |
|
|
|
|
|
|
|
|
|
|
|
|
|
лози. Існує три типи рецепторів гормону. Один |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
із них зв’язує як паратгормон, так і пептид, по- |
|
|
|
— недостатності гіпофіза (вторинний гіпоти- |
|
реоз) чи гіпоталамуса (третинний гіпотиреоз); |
дібний до паратгормону, і через GS активує аде- |
|
|
|
— деяких рідких станах, таких як недостат- |
нілатциклазу. Цей рецептор також може за до- |
|
ність йоду і спричинена лікарськими препарата- |
помогою білка G активувати фосфатидилінози- |
|
q |
|
толову систему. Білок з активністю паратгормо- |
|
ми недостатність щитоподібної залози. |
ну утворюється в багатьох тканинах. У хрящах, |
|
|
|
При дефіциті гормонів щитоподібної залози |
|
утворення тепла і використання кисню зменшу- |
які розвиваються, він стимулює проліферацію |
|
ються, а метаболіти окиснюються повільніше, у |
хондроцитів і пригнічує їх мінералізацію. |
|
результаті чого маса тіла збільшується (гіпоти- |
Механізм дії паратгормону |
|
реоз). Виникають набряки, зумовлені збільшен- |
|
1. Його головна функція — мобілізація каль- |
|
ням гідрофільності тканин (слизовий набряк). |
|
Гіпотиреоз, який виявляється від самого наро- |
цію з кісток і збільшення екскреції фосфату з се- |