Материал: Біохімічна відповідь олігодендроцитів до ішемічного стану
Експериментальні дані демонструють
розподіл загального протеїну (ЗП) у цитозольних фракціях, отриманих із різних
відділів головного мозку щурів (рис. 3. 3). У контрольній групі тварин
показники ЗП наступні: мозочок - 1,501±0,186 мг/мл, кора - 1,16±0,18мг/мл,
таламус - 1,51±0,16 мг/мл і гіпокамп - 2,29±0,18 мг/мл відповідно. Для
ішемічної групи показники інші: мозочок - 1,42±0,22 мг/мл, кора - 1,77±0,19
мг/мл, таламус - 1,09±0,16 мг/мл та гіпокамп - 1,53±0,15 мг/мл відповідно.
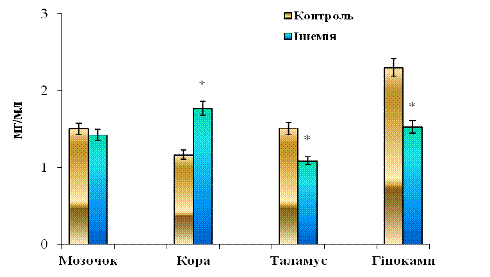
Рис. 3.3 Вміст загального протеїну (ЗП) у цитозольних фракціях, отриманих із різних відділів головного мозку щурів: n = 6; *р<0,05 відносно контролю
Отримані нами результати визначають тенденцію до зменшення рівня загального протеїну (ЗП) у цитозольних фракціях, отриманих з таламусу (на 27,8% у порівнянні з контролем) та гіпокампу (на 33,5% відносно контролю) на відміну від кори великих півкуль, де спостерігається видиме підвищення цього показника у 1,52 рази (на 52,1%). У мозочку не спостерігається достовірних змін у розподілі ЗП, тобто ізадрин та пітуїтрин мало впливають на розподіл ЗП у цитозольній фракції мозочка.
На основі вищезазначеного можна припустити, що за умов глобальної ішемії мозку у таламусі та гіпокампі спостерігається суттєве зниження активності цитозольних ферментів, які відповідають за синтез білкових молекул. Згідно з дослідженнями у гіпокампі поряд зі значним зниженням білкового пулу у цитоплазмі спостерігається активація особливих білків-«протекторів» - сестринів (особливо, секстрину-2). Вони захищають нейрони гіпокампу від ішеміє-індукованого апоптозу шляхом модуляції рибосомального протеїну S6. Доведено, що експресія сестринів [70] індукується ішемією.
У корі стрімке підвищення вмісту загального протеїну, спостерігається, можливо, у зв'язку з компенсаторними реакціями, які «вмикаються» за умов недостатньої концентрації кисню та впливу речовин з катехоламіноподібною активністю, зокрема, ізопреналіну (ізадрину), дофаміну, добутаміну, сальбутамолу. У великих концентраціях катехоламіни стимулюють катаболізм білків [71], внаслідок чого вміст продуктів білкового обміну зростає. У фізіологічних дозах чинять стимулюючий вплив на експресію білків [72].
При ішемії при короткотривалому введенні препаратів останні мало впливають на білковий обмін у мозочку тварин обох груп. Це може свідчити про відносну резистентність останнього до дії катехоламінів, проте ще не свідчить про виняткову стійкість мозочка до ішемічного впливу. Можливо, за максимальних доз та тривалої дії препаратів у мозочку спостерігатимуться певні достовірні варіації розподілу загального протеїну у цитозольної фракції.
У нашому дослідженні показано також розподіл загального протеїну у різних відділах мозку щурів при введенні терапевтичних препаратів - Корвітину, альфа-кетоглутарату (α-КГ) (рис. 3.4-3.7), які широко використовуються при лікуванні церебральної ішемії, ішемічної хвороби серця, інсульту та інших серцево-судинних захворювань.
На рис. 3. 4. показано вміст ЗП у
цитозольних фракціях, отриманих з мозочка головного мозку щурів.
Як зазначено вище, у контрольній та ішемічній групах тварин не виявлено
достовірних змін у концентрації ЗП.
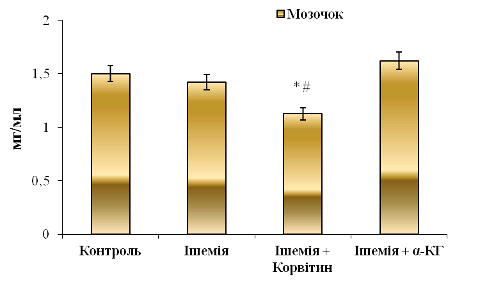
Рис. 3.4 Вміст загального протеїну (ЗП) у
цитозольних фракціях, отриманих з мозочка головного мозку щурів: n = 6;
*р<0,05 відносно контролю; # р<0,05 відносно ішемії
При 5-денному введенні Корвітину та 6-денному введенні альфа-кетоглутарату показники ЗП складають відповідно 1,13±0,19 мг/мл та 1,62±0,10 мг/мл відповідно. При порівнянні з ішемічною групою тварин з рис. 3. 4 видно, що Корвітин знижує вміст ЗП в 1,3 рази (на 20,9 %). При порівнянні з контролем цей показник є нижчим на 24,9%. Такі співвідношення свідчать про те, що препарат здійснює відчутний вплив на клітини мозочка, оскільки знижує вміст продуктів катаболізму білків, знижує стійкість клітин до низьких концентрацій кисню і, в наших дослідженнях, не сприяє проявляє регенеративної дії на нервову тканину мозочка після генералізованої ішемії.
Альфа-кетоглутарат (α-КГ)
виявляє протилежні ефекти: збільшує вміст загального протеїну на 13,9% у
порівнянні з тваринами ішемічної групи та на 8,1% по відношенню до контролю.
Діючою речовиною препарату є 2-оксоглутарова кислота - важливий метаболіт циклу
Кребса, який приймає участь в енергетичних процесах клітини та є попередником
для синтезу таких амінокислот як глутамінова кислота і глутамін. На відміну від
Корвітину препарат підвищує інтенсивність білкового синтезу в уражених ішемією
ділянках мозку, результатом чого є зростання концентрації ЗП у цитозолі. У той
же час препарат чинить слабкий вплив на процеси ПОЛ, менш активно зв'язує
вільнорадикальні форми кисню, які є індуктором ішеміє-залежного апоптозу
уражених клітин. Порівнюючи ЗП у групах тварин з Корвітином та альфа-кетоглутаратом,
бачимо, що в першому випадку вміст білка є нижчим у 1,44 рази (на 43,9%). Це
свідчить про сильніший вплив Корвітину на білковий обмін, причому Корвітин
пригнічує, а α-КГ
- активує білковий синтез. Корвітин знижує вміст токсичних продуктів білкового
обміну, які негативно впливають на життєдіяльність клітин, проте не відновлює
порушені ішемією ділянки у мозочку. α-КГ,
навпаки, сприяє регенерації ушкоджених тканин, і, в підсумку, виявляє набагато
кращий терапевтичний ефект, ніж Корвітин.
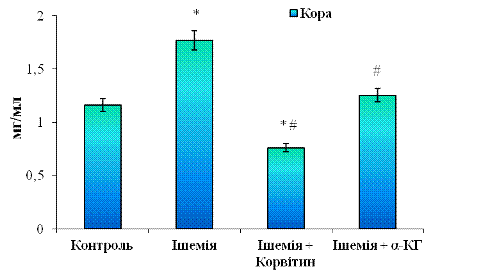
Рис. 3.5 Вміст загального протеїну (ЗП) у
цитозольних фракціях, отриманих з кори великих півкуль мозку щурів: n = 6;
*р<0,05 відносно контролю; # р<0,05 відносно ішемії
Для кори великих півкуль, на відміну від мозочка та інших відділів головного мозку, характерне підвищення рівня ЗП, що вказує на адаптацію клітин в осередкових вогнищах ішемії (див. рис. 3. 5). Цей показник підвищується у 1,52 рази (на 52,1%) відносно контрольної групи.
При 5-денному лікуванні Корвітином концентрація ЗП знижується в 2,3 рази (на 56,9%) при порівнянні з ішемічною групою і становить 0,76±0,11 мг/мл. У випадку α-КГ цей показник складає 1,25±0,14 мг/мл і знижується лише в 1,4 рази (на 29,2%) відносно ішемічної групи. Подібні ефекти є наслідком виснаження метаболічних ресурсів нейронів кори, оскільки, як видно з рис. 3. 5, короткотривала дія ізадрину та пітуїтрину посилює експресію білків.
Схожа картина має місце, якщо порівнювати розподіл ЗП при корвітиновій терапії з контрольною групою. Так, показник білка при введенні Корвітину є нижчим на 34,5%, ніж у нормі (контроль), що можна пояснити білковим виснаженням кори. При введенні α-КГ вміст білка підвищується і є трохи вищим, ніж контроль (приблизно на 7,7%), і це є результатом регенеративних процесів у корі великих півкуль. Якщо ж співставити дані обох препаратів, то вміст ЗП у корі за умов α-КГ-терапії перевищує більш ніж на 50% такий показник для Корвітину. Отже, α-КГ у цьому сприяє покращенню функціонального стану кори і чинить лабільніший вплив на відновлення тканини, аніж Корвітин.
Дані для таламусу представлені на рис. 3. 6. Як згадувалося раніше, у цьому відділі мозку проявляється тенденція до зменшення рівня загального протеїну (ЗП) у цитозольних фракціях на 27,8% при порівнянні з нормою.
Згідно літературних даних [65] відомо, що окремі вогнища ішемії за умов тривалого впливу ішеміє-індукуючих факторів можуть розвинутися у таламічні інфарктів, які є одними з найнебезпечніших патологій головного мозку. Вони виникають внаслідок ураження глибоких гілок таламо-субталамічних артерій, що відходять від проксимального відділу задньої мозкової артерії. Оклюзія їх супроводжується пригніченням свідомості, парезом погляду догори, нейропсихологічними порушеннями, розладами пам'яті (антероградна або ретроградна амнезія) тощо.
При лікуванні Корвітином (див. рис.
3. 6) не виявлено достовірних змін у розподілі білка у таламусі відносно
ішемічної групи. Показник ЗП при цьому склав 1,11±0,14 мг/мл. Можливо, Корвітин
не здійснив помітного впливу на функціональний стан клітин таламусу, що може
спричинитися зменшенням толерантності таламусу до препарату. Відносно норми
(контролю) показник ЗП при корвітиновій терапії є нижчим приблизно на 26,3%.
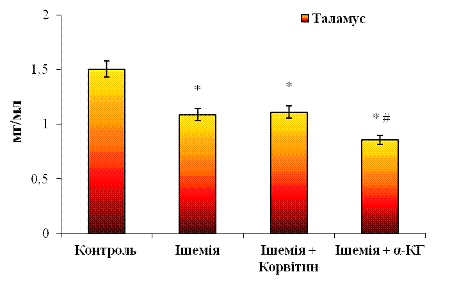
Рис. 3.6 Вміст загального протеїну
(ЗП) у цитозольних фракціях, отриманих з таламусу мозку щурів:
n = 6; *р<0,05 відносно контролю; # р<0,05 відносно ішемії
За умов введення α-КГ концентрація білка у цитозольних фракціях таламусу знизилася в 1,3 рази (на 22,8%) порівняно з ішемією. Звідси бачимо, що клітини таламусу більш чутливі до цього препарату, ніж до Корвітину. Відносно контролю показник є зниженим на 43,1%, тобто α-КГ пригнічує білковий синтез. Після завершення лікування можливе відновлення вмісту ЗП до нормального рівня.
Якщо ж співставити показники білка для Корвітину та α-КГ, бачимо, що в другому випадку вміст ЗП знижений на 22,8% відносно корвітинової групи, тобто α-КГ чинить сильніший вплив на уражені клітини таламусу на відміну від Корвітину. Однак не можна не враховувати той факт, що зниження білка може відбуватися і в нормальних клітинах, розташованих поблизу від вогнищ ішемії.
У цитозольних фракціях гіпокампу, згідно з рис. 3. 7, наявна тенденція до падіння рівня ЗП в усіх досліджуваних групах. Показники для контролю, ішемічної групи, при введенні Корвітину та α-КГ склали 2,23±0,18 мг/мл, 1,53±0,15 мг/мл, 1,26±0,13 мг/мл та 1,08±0,18 мг/мл відповідно.
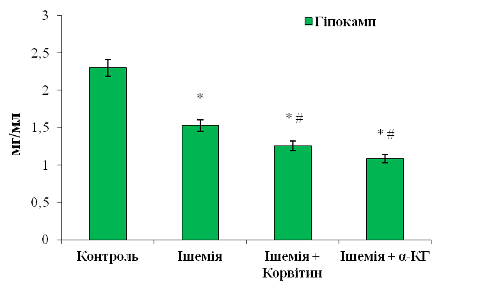
Рис. 3.7 Вміст загального протеїну
(ЗП) у цитозольних фракціях, отриманих з гіпокампу великих півкуль щурів: n
= 6; *р<0,05 відносно контролю; # р<0,05 відносно ішемії
З рис. 3. 7 видно, що у третій і
четвертій групах тварин при введенні препаратів концентрація ЗП суттєво
зменшується по відношенню, як до ішемії, так і до контролю. Для Корвітину
відмічено 1,2-разове зниження (на 17,7%) у порівнянні з ішемічною групою та
1,5-разове зниження (на 33,5%) відносно контролю. Для α-КГ:
1,4-кратне зниження (на 30%) відносно ішемії та 2-кратне (на 52,9%) - відносно
контролю. З цих співвідношень бачимо, що на відміну від інших відділів мозку у
гіпокампі за умов впливу катехоламінів з наступним введенням нейропротекторів
пригнічується пластичний обмін білків. Тому важливий акцент слід зробити на
вивчення механізму реалізації цього впливу та здійснити пошук нових нейропротекторних
препаратів, які чинили менший побічний вплив на ЦНС.
3.4
Основний білок мієліну за умов ішемії
За умов глобальної ішемії вміст
основного білка мієліну є чутливим до впливу ішеміє-індукуючих факторів. Проте
ступінь чутливості є неоднаковим для різних відділів головного мозку. На рис.
3. 8. показано динаміку білка у мозочку, корі, таламусі та гіпокампі
піддослідних тварин.
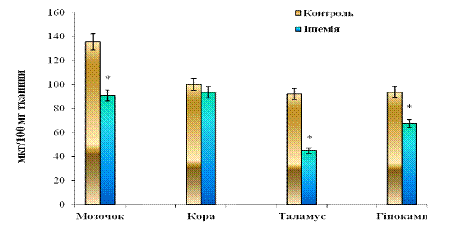
Рис. 3.8 Вміст основного білка мієліну (ОБМ) у
різних відділах головного
мозку щурів: n = 6; *р<0,05 відносно
контролю
Згідно з даними рис. 3. 8 концентрація ОБМ у контрольній групі щурів складають: мозочок - 135,54±0,01, кора - 100,13±0,05, таламус - 92,26±0,01 та гіпокамп - 93,73±0,09 мкг/100 мг тканини відповідно. Для ішемії показники наступні: мозочок - 90,78±0,02, кора - 93,57±0,06, таламус - 45,01±0,02 та гіпокамп - 67,50±0,03 мкг/100 мг тканини відповідно. Звідси видно, що у мозочку, таламусі та гіпокампі спостерігається тенденція до зниження ОБМ у ішемічній групі тварин при порівнянні з контролем. У корі великих півкуль зміни у концентрації білка не є достовірними.
У мозочку при глобальній ішемії мозку наявне падіння білка у 1,49 разів (на 33%) при порівнянні з контролем. У таламусі зниження вмісту ОБМ більш значне, тобто концентрація білка зменшується у 2-3 рази (на 50-55%) відносно контролю. Для гіпокампу продемонстровано 1,39-разове зниження концентрації ОБМ (на 30%) при порівнянні з контрольною групою.
При введенні ізадрину та пітуїтрину (препарати, які індукують розвиток глобальної ішемії) експресія білка у таламусі є найбільш уразливою при ішемії. При цьому має місце демієлінізація нервових волокон, наслідком якої є зменшення концентрації білка. Чутливість мозочка та гіпокампу знаходиться приблизно на одному рівні. Для кори великих півкуль, навпаки, не показано достовірних змін вмісту білка, що свідчить про високу резистентність неокортексу до дії хімічних ішеміє-індукуючих агентів. Рівень ОБМ тут залишається відносно стабільним.
Дослідженнями [35] та [41] продемонстровано різке зниження рівня ОБМ та білкової мРНК, характерне для початкових етапів ішемічного ушкодження мозку гризунів (церебральної ішемії), викликаного високими концентраціями адреналіну (при підшкірному або внутрішньочеревному введенні) чи механічним шляхом (при оклюзії серединної мозкової артерії). Ішемічно-гіпоксичний стан мозку індукує загибель олігодендрогліальних клітин з наступною демієлінізацією аксонів. При цьому порушується структура гемато-енцефалічного бар'єру, що, у свою чергу, призводить до вивільнення ОБМ у кров [42]. Згідно з дослідженнями [43] мінімальні показники протеїну виявлені на першу-другу ішемічну добу впродовж терміну ішемічної експозиції. Так, показано, що експресія ОБМ мРНК та самого білка суттєво зменшується на 23-24-ту годину церебрального ішемічного ушкодження в ішемізованих ділянках, проте підсилюється в периішемічних зонах (зонах, розташованих поряд з ішемізованою ділянкою). На 7-му добу спостерігається стійке підвищення вмісту ОБМ практично у всіх відділах мозку, що свідчить про поступову регенерацію мієлінових оболонок [43]. Проте, за нашими дослідженнями, рівень ОБМ не відновлюється до свого нормального початкового рівня при порівнянні з контрольною групою тварин. При введенні нейропротекторних препаратів - Корвітину та альфа-кетоглутарату фактично не спостерігається відновлення концентрації ОБМ до нормального рівня. Результати, отримані для різних відділів головного мозку щурів при лікуванні вищезазначеними препаратами, представлені на рис. 3. 9, 3. 10, 3. 11 та 3. 12. Група тварин, яку протягом 6-ти днів лікували Корвітином, на рис. позначена «Ішемія + К»; тих тварин, яких лікували альфа-кетоглутаратом, позначено «Ішемія + α-КГ».
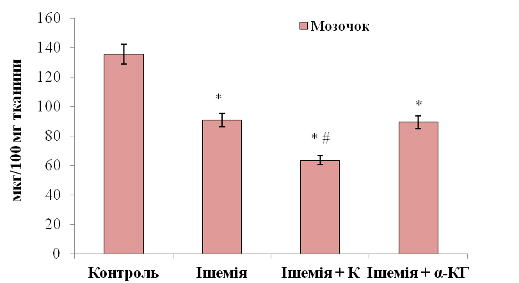
Рис. 3. 9. Вміст основного білка мієліну (ОБМ) у
мозочку щурів: n = 6; *р<0,05
відносно контролю; # р<0,05 відносно ішемії
При введенні нейропротекторних препаратів показники основного білка мієліну у мозочку такі: при корвітиновій терапії - 63,56±0,01 мкг/100 мг тканини, при терапії альфа-кетоглутаратом - 89,31±0,01 мкг/100 мг тканини (див. рис. 3. 9). При порівнянні з ішемічною групою тварин при введенні Корвітину показники ОБМ знизилися у 1,43 рази (на 30%), при порівнянні з контролем - у 2 рази (на 53,1%). Це може вказувати на супутній токсичний вплив препарату на клітини мозочку. При введенні альфа-кетоглутарату не виявлено достовірних варіацій білка відносно ішемії, проте відносно контрольної групи вміст ОБМ є нижчим на 34,1%. Звідси видно, що препарати не відновлюють білу речовину мозочка у постішемічний період.
Для кори великих півкуль (рис. 3. 10) маємо наступний розподіл білка: 68,81±0,01 мкг/100 мг тканини (при введенні Корвітину) та 81,85±0,01 мкг/100 мг тканини (при введенні альфа-кетоглутарату). При порівнянні з ішемічною групою тварин: Корвітин знизив вміст ОБМ на 26,5%, альфа-кетоглутарат - на 12,5%. Відносно контролю: Корвітин - на 31,3%, альфа-кетоглутарат - на 18,3%.
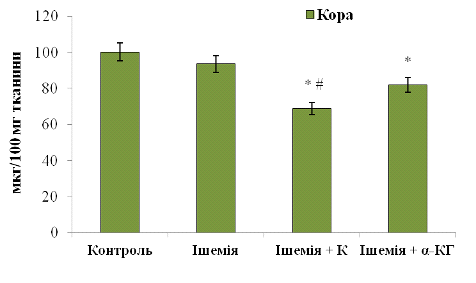
Рис. 3.10 Вміст основного білка
мієліну (ОБМ) у корі великих півкуль мозку щурів: n
= 6; *р<0,05 відносно контролю; # р<0,05 відносно ішемії
Для таламусу (рис. 3. 11) показники
ОБМ наступні: 47,41±0,07 мкг/100 мг тканини (у групі «Ішемія + Корвітин») та
57,82±0,07 мкг/100 мг тканини. При порівнянні концентрації ОБМ у ішемічній та
корвітиновій групах тварин не спостерігається вірогідних змін; при введенні
альфа-кетоглутарату наявне невелике підвищення вмісту білка (приблизно на
18-20%), однак це свідчить про незначний ступінь регенерації ушкоджених ішемією
тканин таламусу. Відносно контролю (норми) показники білка знижені на 48,6%
(Корвітин) та на 37,3% (альфа-кетоглутарат).
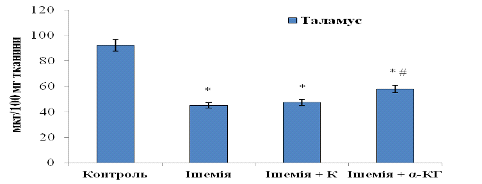
Рис. 3. 11. Вміст основного білка мієліну (ОБМ) у таламусі щурів: n = 6; *р<0,05 відносно контролю; # р<0,05 відносно ішемії
У випадку гіпокампу (рис. 3. 12)
вміст ОБМ виглядає так: 60,45±0,02 мкг/100 мг тканини (група «Ішемія +
Корвітин») та 59,30±0,04 мкг/100 мг тканини (група «Ішемія + α-КГ»).
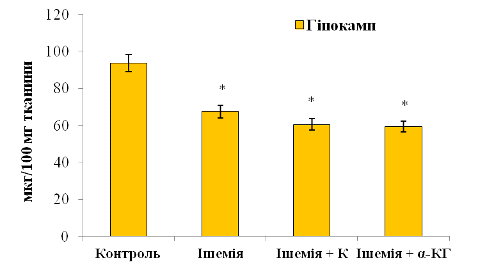
Рис. 3.12. Вміст основного білка мієліну (ОБМ) у
гіпокампі щурів: n
= 6; *р<0,05 відносно контролю; # р<0,05 відносно ішемії
Відносно ішемії не виявлено суттєвих варіацій білка. При порівнянні з контрольною групою показники знижені на 35,5% (Корвітин) і 36,7% (альфа-кетоглутарат) відповідно. Видно, що у гіпокампі спостерігається схожа з іншими відділами мозку тенденція до зменшення кількості білка у цитозольних фракціях по відношенню до норми. Це вказує на виснаження пластичного балансу уражених олігодендроглії, ураженою ішемією. Причому видно, що препарати чинять незначний терапевтичний ефект, при цьому знижуючи метаболізм специфічних протеїнів ЦНС.