Материал: А27483 Красникова ЛВ Кострова ИЕ Машкин ДВ Микробиология производства хлеба кондит и макарон изделий
Оценка дрожжей по показателю мальтазной активности
|
Мальтазная активность, мин |
Оценка дрожжей |
|
85–100 |
Отличные |
|
101–110 |
Хорошие |
|
111–160 |
Удовлетворительные |
|
Свыше 160 |
Плохие |
О с м о ч у в с т в и т е л ь н о с т ь дрожжей, т. е. чувствительность к определенной концентрации сахара и хлорида натрия, определяют по разнице во времени, которое требуется на выделение 10 мл СО2 навесками дрожжей в тесте с добавлением хлорида натрия и без него.
На кафедре технологии хлебопекарного производства МГУПП совместно с другими научными коллективами проведен комплекс исследований по разработке методов оценки свойств дрожжей и введено понятие «биотехнологические свойства дрожжей».
В понятие «биотехнологические свойства дрожжей» входит основной критерий оптимальности процесса брожения – максимальная скорость газообразования в тестовых полуфабрикатах, что обусловливает общую продолжительность технологического процесса и формирование качественных показателей полуфабрикатов и готовой продукции.
В динамике газообразования дрожжей имеют место два периода. В первый период дрожжевые клетки образуют диоксид углерода при сбраживании моносахаров (глюкозы и фруктозы). При этом глюкоза сбраживается непосредственно, а фруктоза – после изомеризации ее индуцируемым ферментом фруктоизомеразой. Присутствующая в среде сахароза расщепляется под действием β-фруктофуранози-дазы дрожжей на глюкозу и фруктозу. Затем скорость газообразования снижается и происходит адаптация дрожжей к мальтозе. В этот период дрожжи начинают синтезировать два фермента: мальтопермеазу, локализованную на поверхности цитоплазматической мембраны (функция этого фермента – транспортировка мальтозы, полученной в результате расщепления крахмала a- и b-амилазами муки, в дрожжевую клетку), и a-глюкозидазу, расположенную внутри клетки. Последний фермент расщепляет мальтозу, доставленную в клетку мальтопермеазой, на две молекулы глюкозы, которая затем сбраживается дрожжами при участии зимазного комплекса. Во втором периоде скорость газообразования достигает максимального значения.
Отсюда следует, что созревание полуфабрикатов необходимо доводить до оптимума. Качество дрожжей предложено оценивать по бродильной активности, выражающейся объемом диоксида углерода (в сантиметрах кубических на 1 г СВ дрожжей при давлении 760 мм рт. ст.), выделившегося при брожении до достижения максимальной скорости газообразования.
Оценка бродильной активности дрожжей позволяет, наряду с подъемной силой, учитывать и их мальтазную активность, что дает более полную картину биотехнологических свойств используемых дрожжей.
1.1.5. Штаммы (расы) хлебопекарных дрожжей
Штаммами (расами), как уже указывалось, называют микроорганизмы одного вида, выделенные из разных источников или из одного источника, но в разное время и различающиеся между собой некоторыми второстепенными признаками.
Для хлебопекарного производства рекомендованы следующие расы и штаммы дрожжей.
Раса Томская-7 (Т-7). Клетки круглые или слегка овальные, довольно мелкие – (6–8)(5–6) мкм. Обладают хорошей зимазной, но слабой мальтазной активностью (более 160 мин). Культура устойчива к изменениям состава мелассы, однако требовательна к содержанию витаминов в среде. Прессованные дрожжи, полученные на этой расе, являются стойкими при хранении.
Раса Одесская-14 (О-14). Клетки овальные или эллипсовидные размером (7–11)(6–8) мкм. Культура отличается высокой скоростью генерации; дает выход дрожжей выше, чем расы Томская-7 и ЛДБ-Х1. Дрожжи этой расы стойки при хранении в прессованном виде и устойчивы к высушиванию, однако они требовательны к составу питательной среды, особенно к наличию в ней ростовых веществ.
Раса Киевская-21 (К-21). Клетки овальные или круглые размером (6–10)(5–6,4) мкм. Культура устойчива к ингибиторам роста, нетребовательна к ростовым веществам, хорошо переносит высушивание, обладает хорошей зимазной и мальтазной активностью.
Штамм Л-441. Выведен в Ленинградском отделении ГосНИИХП путем отбора на основе естественной изменчивости дрожжей расы Одесская-14. Клетки овальные, крупные, размером (8–14)(6–9) мкм. Характеризуется высокой продуктивностью, полностью сбраживает раффинозу, устойчив к вредным примесям и микробной обсемененности мелассы. Имеет высокую удельную скорость роста и обеспечивает хорошие хлебопекарные свойства товарных дрожжей (зимазная активность – 45 мин, мальтазная – 120 мин). При температуре 35 ºС хранится свыше 96 ч.
Штамм Я. Выведен на Янгиюльском дрожжевом заводе из производственной чистой культуры расы О-14 путем направленного отбора. Обладает высокой скоростью генерации и устойчивостью к повышенным температурам выращивания (37–38 ºС), что немаловажно для предприятий южных регионов. Клетки крупные, округлой формы. Подъемная сила составляет 40–47 мин, зимазная активность – 32–44 мин.
Штаммы Л-1, Л-2, Л-3, ЛК-14 селекционированы в Ленинградской лаборатории ВНИИПБТ. Имеют клетки средних и крупных размеров. Характеризуются высокой продуктивностью, способны конкурировать с посторонней микрофлорой, устойчивы к повышенной температуре (до 40 ºС). Штаммы накапливают большое количество трегалозы (Л-1 и Л-2 – 10–12 %, остальные – 14–17 %) и хорошо переносят высушивание, сохраняя исходную ферментативную активность. Зимазная активность прессованных и сушеных дрожжей для всех штаммов составляет 45–56 мин; мальтазная активность штаммов: Л-2 – 60–80 мин; Л-1 – 100–110 мин; Л-3 – 120–135 мин; ЛВ-7 и ЛК-14 – 130–180 мин.
Штамм 616 накапливает значительное количество трегалозы (до 16,2 % на СВ), что позволяет рекомендовать его для производства сушеных дрожжей. Зимазная активность штамма – 55 мин, ά-глюко-зидазная активность – 67 мин, осмочувствительность – 5–10 мин.
Гибридные расы 176, 196-6 и 262 имеют крупные клетки, обладают высокой устойчивостью к составу мелассных сред; их зимазная активность составляет 42–57 мин, мальтазная активность – 65–75 мин. Раса 262 рекомендована для получения сушеных дрожжей (содержание трегалозы составляет 8,7 % на СВ).
Штамм ЛВ-7 используется для производства прессованных и сушеных дрожжей. Он обладает высокой устойчивостью к составу мелассных сред и посторонней микрофлоре, контаминирующей дрожжевое производство; характеризуется повышенной продуктивностью и способностью накапливать значительное количество трегалозы (в два раза выше, чем аналоги).
Штамм 722 имеет хорошие показатели по мальтазной активности – 54 мин, зимазной активности – 43 мин, подъемной силе – 46 мин и осмочувствительности – 5–10 мин. Этот штамм используют для производства сушеных дрожжей.
Штамм 739 отличается высокой продуктивностью и ферментативной активностью. Дрожжи этого штамма полностью сбраживают глюкозу, фруктозу, сахарозу, мальтозу, раффинозу и галактозу. Имеют зимазную активность – 54 мин, мальтазную активность – 61 мин, подъемную силу – 56 мин.
Штамм 93, обладающий высокой продуктивностью и активным комплексом ферментов, используют для производства сушеных дрожжей. Его зимазная активность составляет 45 мин, мальтазная – 53 мин, подъемная сила – 45 мин.
Штамм 512 – гибридный, получен путем скрещивания штамма расы ХII и штамма Saccharomyces diastaticus. Гибрид является триплоидом, он характеризуется повышенным синтезом витаминов: эргостерина, тиамина, рибофлавина, пиридоксина, ниацина. Имеет осмочувствительность 14 мин.
Штамм 5 является гибридом от скрещивания двух штаммов дрожжей – «Яблочного 3», применяемого при сбраживании яблочного сока, и штамма 722. Полученный штамм характеризуется высокой бродильной активностью (табл. 1.2).
Штамм 69 получен путем скрещивания расы дрожжей «Джамбулская-60» и штамма 10, выделенного из французских сушеных дрожжей. Для штамма 69 характерны высокая скорость роста, зимазная активность – 45 мин, мальтазная – 90 мин. Штамм устойчив к повышенным температурам – 40–45 ºС.
Штамм V-503 – мутант, полученный путем воздействия лазерного света на штамм 73. По ряду показателей: выходу биомассы, подъемной силе, зимазной и мальтазной активности – мутантный штамм превосходит родительский. Штамм V-503 имеет хорошие хлебопекарный свойства: подъемная сила составляет 19,8 мин; ά-глюкози-дазная и зимазная активность составляют соответственно 25 и 35 мин; стойкость – 100 ч при 35 ºС
Хлебопекарные свойства некоторых штаммов дрожжей вида Saccharomyces cerevisiaе приведены в табл. 1.2.
Таблица 1.2
Хлебопекарные свойства некоторых рас и штаммов дрожжей
|
Показатель, мин |
Т-7 |
К-21 |
О-14 |
Л-1 |
616 |
ЛВ-7 |
5 |
|
Зимазная активность |
55 |
60 |
45 |
45 |
55 |
45 |
85 |
|
Мальтазная активность |
160 |
100 |
95 |
110 |
67 |
120 |
95 |
|
Осмочувстви- тельность |
12 |
10 |
10 |
18 |
5–10 |
6–10 |
15 |
Среди дрожжей вида Saccharomyces minor в хлебопечении используют штаммы 12/17, 7 и «Чернореченский».
Штамм 12/17. Клетки овальной формы размером (2,9–3,0) (4,0–4,2) мкм. Штамм имеет подъемную силу (по шарику) 19–21 мин, выдерживает кислотность до 14 град.
Штамм 7. Клетки мелкие круглые или слегка овальные размером 2,62,2 мкм. Подъемная сила – 17–19 мин, может размножаться при кислотности до 16 град.
Штамм «Чернореченский». Клетки овально-округлой формы размером 3,8–3,2 мкм. Подъемная сила 19–20 мин. Штамм способен размножаться при кислотности среды 16–18 град.
1.1.6. Спиртовое брожение, осуществляемое дрожжами
Размножение дрожжей в тесте происходит в анаэробных условиях, в результате чего они получают энергию за счет спиртового брожения. Образующийся при этом брожении диоксид углерода разрыхляет тесто.
Процесс спиртового брожения суммарно можно выразить следующим уравнением:
С6Н12О6 + 2 Фн + 2 АДФ → 2 СО2 + 2 С2Н5ОН +2 АТФ + 2 Н2О
глюкоза диоксид этанол
углерода
Спиртовое брожение протекает по фруктозодифосфатному пути (гликолитический путь Эмбдена–Мейергофа–Парнаса) (рис. 1.5).
Первая реакция процесса брожения – фосфорилирование глюкозы при участии АТФ с образованием глюкозо-6-фосфата. Затем молекула глюкозо-6-фосфата изомеризуется в молекулу фруктозо-6-фосфата, которая, в свою очередь, фосфорилируется в положении 1. Образовавшийся при этом фруктозо-1,6-дифосфат расщепляется под действием фермента альдолазы (ключевого фермента гликолитического пути) на две триозы – диоксиацетонфосфат и 3-фосфоглицери-новый альдегид (3-ФГА). Диоксиацетонфосфат превращается в 3-ФГА в результате реакции, катализируемой триозофосфатизомеразой. На этом заканчивается первая стадия гликолитического пути, характеризующаяся активированием одной молекулы глюкозы с затратой энергии в виде двух молекул АТФ. На второй стадии брожения происходит окисление 3-ФГА до 1,3-дифосфоглицериновой кислоты при участии НАД-зависимой 3-фосфоглицеральдегиддегидрогеназы. Далее 1,3-дифосфоглицериновая кислота взаимодействует с АДФ, отдавая фосфатную группу, что приводит к синтезу АТФ. На этом этапе имеет место первое субстратное фосфорилирование, т. е. высвобождение энергии и запасание ее в виде АТФ. В следующей реакции гликолиза фосфатная группа 3-фосфоглицериновой кислоты переносится из третьего положения во второе.
Затем происходит реакция дегидратации 2-фосфоглицериновой кислоты, катализируемая ферментом енолазой, приводящая к образованию фосфоенолпирувата. От молекулы фосфоенолпирувата фосфатная группа переносится на АДФ с помощью фермента пируваткиназы, что приводит к образованию АТФ и пирувата. Это вторая реакция субстратного фосфорилирования.
Таким образом, в результате гликолиза из одного моля глюкозы образуются два моля пирувата, от которого с помощью фермента пируватдекарбоксилазы – ключевого фермента спиртового брожения – отщепляется диоксид углерода. Образовавшийся при этом ацетальдегид становится акцептором водорода окисляемого субстрата и восстанавливается до этанола с участием НАД+-зависимой алкогольдегидрогеназы. Биологическое значение последней реакции заключается в окислении восстановленного НАДН+Н+, образовавшегося ранее в ходе реакции превращения 3-фосфоглицеринового альдегида в 1,3-ди-фосфоглицериновую кислоту. Данный путь включает как использование АТФ, так и образование АТФ, давая чистый выход 2 моля АТФ на каждый моль сбраживаемой глюкозы.
К. Нойберг обнаружил, что в зависимости от условий при спиртовом брожении могут образовываться нехарактерные для этого процесса продукты. Так, если к дрожжам, сбраживающим глюкозу, добавить бисульфит (NaHSO3), то последний образует комплекс с ацетальдегидом, в связи с чем он больше не может выполнять роль акцептора электронов. Вследствие этого электроны от восстановленного НАДН передаются на диоксиацетонфосфат. Он восстанавливается до глицерол-3-фосфата и дефосфорилируется с образованием глицерина, как это показано на схеме (см. рис. 1.5). Это модифицированное брожение называют глицериновой формой спиртового брожения.
ГЛЮКОЗА
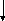 АТФ
АТФ
1 АДФ
Глюкозо-6-фосфат
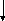 2
2
Фруктозо-6-фосфат
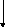 АТФ
АТФ
3 АДФ
Фруктозо-1,6-дифосфат
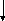
4
 Диоксиацетонфосфат
Глицеральдегид-3-фосфат
Диоксиацетонфосфат
Глицеральдегид-3-фосфат

 5
6
5
6
13 НАДН+Н+ НАД+
НАД+ + Фн НАДН+Н+
Глицерол-3-фосфат 1,3-дифосфоглицериновая
 14
кислота
14
кислота