Материал: Polioxialkanoaty_POA__biorazrushaemye_polimery_dlya_meditsiny
Однако низкая энергетическая эффективность реакции сопровождается большими затратами метанола на образование биомассы и полимера (таблица 2.6). Экономический коэффициент синтеза ПОБ на метаноле составляет 0.18–0.20 г ПОБ/г.
Водород рассматривается как альтернативный субстрат по отношению к сахарам и метанолу, но трудности ферментации на газовом субстрате из-за его взрывоопасности и плохой растворимости, накладывают определенные ограничения на этот субстрат при его рассмотрении применительно к крупнотоннажным процессам.
При росте водородных бактерий рода Alcaligenes на водороде фиксация углекислоты (источника углерода) реализуется в цикле Кальвина со следующими затратами АТФ и восстановительных коэффициентов:
6 СО2 + 16АТФ + 10НАДФН 2ФГК + 10НАДФ+ + 16АДФ + 16Рi
если фосфоглицериновая кислота (ФГК) конвертирует в ПОБ через фосфоэнолпируват, реакция синтеза пирувата и ацетил-КоА выглядит, как:
ПОБn + 2ФГК + НАД + 2АДФ + 2Рi ПОБn + 1 + 2АТФ + 2НАДН + 2CО2. Суммарная реакция может быть представлена, как:
ПОБn + 4СО2 + 14АТФ + 10РАДФН + НАД+ПОБn + 1 + 14АДФ + 14Рi + 10НАДФ+ + НАДН
Эффективность биосинтеза ПОБ на водороде в качестве энергетического субстрата высокая; затраты субстрата на образования продукта – минимальны. Экономический коэффициент получения полиоксиалканоатов на водороде составляет 1. Поэтому освоение этого субстрата может стать перспективным при развитии уровня техники ферментации. К настоящему времени достигнуты достаточно высокие показатели процесса (Tanaka et al., 1995); получен урожай в периодической культуре Alcaligenes eutrophus свыше 90 г/л при внутриклеточной концентрации ПОБ 67.8 %. Оптимизация режима газообеспечения культуры и создание установок с высокими массообменными характеристиками позволили повысить выход полимера в автотрофной культуре водородных бактерий до 80–90 г/л
(Ishizaki et al., 2001).
Таким образом, в качестве возможных субстратов для крупнотоннажного производства ПОА исследуются а) известные субстраты
– смеси водорода и углекислоты, глюкоза, фруктоза, маниока, меласса, лактоза, молочная сыворотка, метанол, тапиока, (Tanaka et al., 1995; Kim et al., 1994b; Ryu et al., 1997; Yamane et al., 1996; Wong and Lee, 1998; Sugimoto et al., 1999) б) новые субстраты: углеводо-
родные кислоты (пентановая, олеиновая, октановая, нонановая,) и
октан (Haazenberg, Witholt, 1997), глюконат (Rodriges et al., 1995),
100
белковые гидролизаты (пептона, казеина) (Borman et al., 1998;), кси-
лоза (Lee, 1998), гидролизаты гемицеллюлоз (Ramsay et al., 1994),
смеси жирных кислот животного и растительного происхождения (Cromwick et al., 1996), отходы производства оливкового и пальмового масла (Tan et al., 1997), а также в) необычные субстраты: метак-
риловая кислота (Lee et al., 1997), триглицериды (Ashby, Foglia, 1998;
Solaiman et al., 1999) и жидкие продукты переработки углей
(Füchtenbusch, Steinbüchel, 1999).
В связи с известной способностью бактерий Pseudomonas и мутантов на их основе синтезировать средне- и длинноцепочечные ПОА, достаточно активно исследуется возможность реализации процесса получения полимеров на жирах и жирных кислотах, в том числе за счет привлечения отходов переработки с/х масличных культур. Так, на олеиновой кислоте возможно получение шестикомпонентных ПОА при общем содержании полимера от 15 до 45 % в проточной культуре P. putida KT2442 в диапазоне D 0.05–0.27 ч–1 (Huijberts, Eggink, 1996). Штамм P. putida PGA1 при культивировании на жирных кислотах, содержащихся в отходах производства пальмового масла, синтезирует шести- и семикомпонентные ПОА c молекулярной массой от 124 000 до 174 000 Da при общем урожае культуры 3.0–8.8 г/л и выходах ПОА от 19 до 37 % (Tan et al., 1997). Трансгенные штаммы Alcaligenes H16 на жиросодержащих средах, получаемых из оливкового, пальмового и кукурузного масел синтезируют до 76–82 % ПОА
(Fukui, Doi, 1998). Pseudomonas oleovorans и мутантные штаммы данного организма с phb-генами из Alcaligenes, а также рекомбинантные штаммы E. coli содержащие гены синтеза ПОА из Ps. oleovorans, при росте на смесях различных жирных, включая насыщенные и ненасыщенные ЖК, 3-гидроксиалкановые кислоты, а также алканы с длиной углеродной цепи от С8 до С18 способны синтезировать многокомпонентные ПОА, содержащие, помимо оксибутирата, оксигексаноат, оксиоктаноат, оксидодеканоат, от следовых включений до 14–80 мол. % для отдельных мономеров. Общий выход ПОА при этом достигает свыше 60 % (Füchtenbusch et al., 2000).
Для получения разнообразных по составу ПОА в качестве возможных источников углерода исследованы самые различные, часто нетрадиционные, субстраты. Среди них – 1,3-бутандиол, обеспечивающий синтез широкого спектра длинноцепочечных ПОА бактериями Pseudomonas sp A33; субстраты, содержащие триглицериды, на которых бактерии Pseudomonas resinovorans и Pseudomonas saccharophila способны синтезировать различные по составу длинноцепочечные ПОА с достаточно высокими выходами, до 44–50 % (к весу сухой биомассы). При этом уровень таких включений, как оксиоктаноат, оксидеканоат, оксидодеканоат достигают 30–40 мол. % (Achby and Foglia, 1998; Solaiman et al., 1999). Проведено исследова-
101
ние октана для продукции полиоксиалканоатов различными культу-
рами Pseudomonas (Ps. oleovorans, Ps. thruringiensis, Ps. Putida) в пе-
риодической культуре, а также в проточных режимах (Hazenberg, Witholt, 1997). Авторами установлено, что возможны достаточно продуктивные процессы синтеза ПОА в периодической и в проточной культурах Pseudomonas oleovorans, до получения урожая биомассы в 40–93 и 11–18 г/л соответственно. Установлена также возможность синтеза ПОА, содержащих в качестве мономеров включения с различной длиной углеродной цепи, от С4 до С14 культурами Pseudomonas (Ps. putida, Ps. oleovorans, Ps. citronellois, Ps .resinovorans) с
использованием жирных кислот и триглицеридов активного ила (Cromwick et al., 1996). Показана применимость для синтеза среднецепочечных ПОА бактериями Psеudomonas putida продуктов омыления масел и жирных кислот из отходов, образуемых при переработке пальмового масла (Tan et al., 1997). Образуемые полимеры в зависимости от жирнокислотного состава субстрата содержат в качестве сополимеров оксикислоты С6, С8, С12 и С14. Сравнительно недавно успешно реализован в промышленном масштабе процесс биосинтеза сополимеров оксибутирата и оксигексаноата бактериями при роста на глюкозе и в качестве индуктора для синтеза среднецепочечных ПОА лауриновой кислоте (Chen et al., 2001).
Выявлено, что мутантные штаммы дрожжей Candida rugosa способны синтезировать D- -оксиизомасляную кислоту при использовании изомасляной кислоты как источника углерода (Kim et al., 1999); концентрация биомассы в культуре составляет до 30 г/л. Полученные в результате селекции бактерии Azotobacter beijerinckii при росте на органических источниках углерода (казеиновый пептон, дрожжевые экстракты и гидролизаты) в сочетании с сахарами (глюкозой или сахарозой) синтезируют полиоксибутират с различной интенсивностью, от единиц до 55 % ПОА к сухому весу клеточной биомассы (Bormann et al., 1998). Alcaligenes eutrophus DSM 11348 на средах с глюкозой, а
также комплексными органическими источниками азота (гидролизаты казеина и смеси аминокислот) в течение короткого ферментационного периода (около 40 часов) синтезируют с различными выходами, включая высокие (свыше 80 %), полиоксибутират. Процесс масштабирован в 300-т литровом аппарате (Bormann et al., 1998).
Рассматриваются также возможности получения полиоксиалканоатов с использованием отходов. Практически неисчерпаемым источником сырья для крупнотоннажного получения полиоксиалканоатов могут стать растительные биомассы, образуемые в огромных количествах ежегодно. Отходы различных сельскохозяйственных культур, содержащие полисахариды различной структуры и состава, могут быть гидролизованы с получением спектра водорастворимых сахаров. Например, показано, что рекомбинантные штаммы E coli,
102
трансформированные генами Al. eutrophus, способны синтезировать полиоксибутират на средах, содержащих ксилозу (20 г/л) с добавками органических форм азота, получаемых из растительных отходов. Среди них – кукурузные и соевые гидролизаты, гидролизаты вегетативной биомассы хлопчатника и др. (Lee, 1998). Гидролизаты целлюлозы, содержащие ксилозу, обеспечивают урожаи и выходы ПОА у бактерий Ps. сepacia, сопоставимые с процессом на фруктозе (Ramsay et al., 1995); продукционная цена полиоксибутирата при этом аналогична процессу на мелассе и в два раза ниже, по сравнению с глюкозой.
В настоящее время все чаще появляются работы, рассматривающие в качестве сырья для ряда биотехнологических производств угли и продукты их переработки (Catcheside, Ralph, 1999; Fakoussa, Hofirichter, 1999). В связи с огромными запасами и относительно низкой стоимостью данный субстрат представляет интерес и для будущих крупнотоннажных производств микробных биопластиков. Такие исследования начаты. Недавно опубликована работа по изучению продукции среднецепочечных полиоксиалканоатов несколькими культурами Pseudomonas на средах, содержащих в качестве источника углерода продукты переработки углей – смеси гуминовых кислот (Füchtenbusch, Steinbüchel, 1999). Установлено, что бактерии Ps. oleovorans способны аккумулировать ПОА, содержащие 3- оксигексаноат, 3-оксидеканоат и 3-оксидодеканоат, а бактерии Rhodobacter rubber – смеси 3-оксибутирата и 3-оксивалерата. Показано, что продукты гидролиза углей, полученные биологическим путем на основе культуры Trichoderma, является для Ps. oleovorans по сравнению с продуктами химического гидролиза углей, более выгодным субстратом. В перспективе возможна организация двух-, трехэтапных биотехнологических процессов, в ходе которых на первых стадиях угли будут трансформироваться до водорастворимых продуктов, например, грибными культурами, а далее, эти субстраты можно будет использовать для синтеза ПОА.
Выполненные сравнительно недавно технико-экономические оценки свидетельствуют о том, что в принципе возможно создание экономически приемлемых крупнотоннажных производств ПОА на различном сырье, включая отходы сельскохозяйственных и про-
мышленных производств (Choi, Lee, 1997; 1999; Hazenberg, Witholt, 1997; Braunegg et al.,1998; Natano et al., 2001).
103
Глава 3. СВОЙСТВА ПОЛИОКСИАЛКАНОАТОВ
3.1. Молекулярная структура и физико-химические свойства ПОА
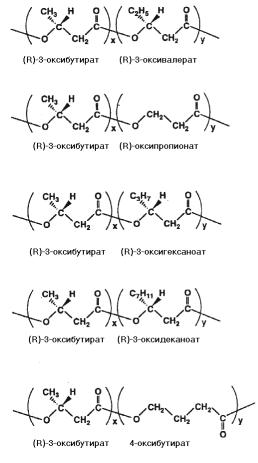
Полиоксиалканоаты, синтезируемые природными и генетически модифицированными организмами, имеют различное химические строение, определяемое соотношением входящих в их состав мономерных единиц, характеризуются различными физикомеханическими свойствами и могут быть использованы для получения различными методами пленок, волокон, нетканых материалов и др. (Amass et al., 1998; Sudesh et al., 2000 и др.). Среди идентифици-
рованных к настоящему времени свыше 100 ПОА (Steinbüchel and Valentin, 1995).
Продуцент
Ralstonia eutropha
Ralstonia eutropha
Alcaligenes latus Aeromonas cavie
Pseudomonas sp.
Ralstonia eutropha
Alcaligenes latus Comamonas acidovorans
Субстрат |
Структура сополимеров |
пропионовая кислота
пентановая кислота 3-оксимасляная кислота
1,5-пентандиол
растительные жиры
сахара
4-оксимасляная кислота
γ-бутуролактон 1,4-бутандиол 1,6-гександиол
Рис. 3.1. Структуры ПОА, содержащих в качестве базового мономера 3-оксибутират и другие мономеры (Sudesh et al., 2000)
104