Материал: 1010-pochvovedenie-2013-7
ВЛИЯНИЕ ТЕМПЕРАТУРЫ НА ИНТЕНСИВНОСТЬ РАЗЛОЖЕНИЯ |
811 |
Лет |
|
|
|
А |
|
|
|
|
40 |
|
|
|
|
|
|
|
|
35 |
|
|
|
|
|
|
|
|
30 |
|
|
|
|
|
|
|
|
25 |
|
|
|
|
|
|
|
|
20 |
|
|
|
|
|
|
|
|
15 |
|
|
|
|
|
|
|
|
10 |
|
|
|
|
|
|
|
|
5 |
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
|
2 |
12 |
22 |
2 |
12 |
22 |
2 |
12 |
22 |
Растительные |
|
С4 – ПОВ |
|
С4 – ПОВ |
||||
|
остатки |
|
контроль |
|
NPK |
|
||
600 |
|
|
|
Б |
|
|
|
|
|
|
|
|
|
|
|
|
|
500 |
|
|
|
|
|
|
|
|
400 |
|
|
|
|
|
|
|
|
300 |
|
|
|
|
|
|
|
|
200 |
|
|
|
|
|
|
|
|
100 |
|
|
|
|
|
|
|
|
0 |
12 |
22 |
2 |
12 |
22 |
2 |
12 |
22 |
2 |
||||||||
|
ПОВ пар |
|
С3 – ПОВ |
|
С3 – ПОВ |
|||
|
|
|
|
контроль |
|
NPK |
|
|
°С
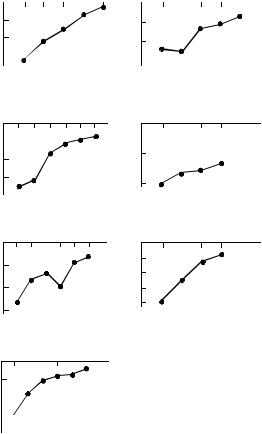
Рис. 7. Средние времена обновления лабильных (А) и устойчивых (Б) пулов ПОВ в зависимости от тем пературы.
кумулятивных потерь СО2 при разложении ПОВ пара, имеют несколько завышенные значения.
Величина Q10 для ПОВ пара в диапазоне темпе ратур 12–22°С составила 3.4, что меньше величи ны 4.3, полученной для С3 ПОВ. Учитывая пря мо пропорциональную зависимость между вели чиной Q10 и устойчивостью пула ПОВ в этом интервале температур (рис. 6), значение темпера турного коэффициента для ПОВ пара свидетель ствует о его меньшей устойчивости по сравнению с С3 ПОВ в почве под растениями кукурузы.
Величины среднего времени обновления рас тительных остатков (рис. 7) изменялись от 2 до 5 лет, С4 ПОВ – от 5 до 35 лет, а С3 ПОВ – от 35 до 500 лет в зависимости от температуры. При та ком соотношении величин MRT очевидно, что наибольший вклад в эмиссию СО2 в полевых условиях будет вносить минерализация лабиль ных пулов (растительных остатков и С4 ПОВ). В окрестностях Воронежа, где расположены иссле дуемые почвы, многолетняя среднегодовая тем пература воздуха составляет 6.5°С [2]. Недавно опубликованные данные свидетельствуют о том, что в период с 1990 по 2010 гг. средняя температу ра в отдельные годы достигала 11.0°С [10]. Данная
экстремальная величина приближается к интер валу 12–22°С, в котором заметно увеличиваются температурные коэффициенты Q10 для констант разложения С3 ПОВ. Таким образом, сохраняю щаяся современная тенденция к потеплению климата может в первую очередь повлиять на ми нерализацию устойчивого пула органического ве щества в исследуемых черноземах. Для того что бы определить, приведет ли это к уменьшению за пасов ПОВ в течение последующих десятилетий, вызвав эмиссию дополнительных количеств СО2 в атмосферу, необходимо в дальнейших исследо ваниях учитывать изменение влажности почвы при потеплении климата.
ЗАКЛЮЧЕНИЕ
Установлено, что практически все изученные показатели цикла углерода в почве – константы разложения органического вещества почвы, ве личина затравочного эффекта, значения темпера турного коэффициента Q10, соотношение между новым (С4) и старым (С3) углеродом в потоке СО2 из почвы – зависят от температуры. Величину за травочного эффекта и соотношение потерь угле рода при разложении лабильного и устойчивого пулов органического вещества почвы в инкуба ционных экспериментах обычно определяют при 22–25°С, не учитывая зависимости этих показа телей от температуры. Закономерности, установ ленные при высокой температуре, вряд ли спра ведливы для полевых условий, поскольку для большинства почв России, расположенной в уме ренном климатическом поясе, температура поч вы редко достигает таких высоких значений. Кро ме того, температурные зависимости не универ сальны, они имеют свои особенности для разных пулов углерода, поэтому определения скоростей различных процессов углеродного цикла как функции температуры в зависимости от типа поч вы и фитоценоза остаются чрезвычайно актуаль ными.
Для более точного описания разложения орга нического вещества почвы недостаточно рас сматривать этот процесс как химическую реак цию распада органического вещества. Тот факт, что корреляция между величиной Q10 и устойчи востью органического вещества, характерная для химической реакции, наблюдалась в исследуе мом черноземе только в ограниченном интервале температур 12–22°С, свидетельствует о необхо димости рассматривать деструкцию органическо го вещества почвы как биохимический процесс. При таком подходе, наряду с определением пото ков углерода, важно оценивать активность клю чевых окислительных и гидролитических фер ментов – катализаторов многостадийного про цесса разложения органического вещества.
ПОЧВОВЕДЕНИЕ № 7 2013
812 |
ЛАРИОНОВА и др. |
СПИСОК ЛИТЕРАТУРЫ
1.Барре П., Эглин Т., Кристенсен Б.Т., Чиайс П., Хо уот С., Каттерер Т., Когут Б., Ван Оорт Ф., Пейлин П., Поултон П.Р., Романенков В., Ченю К. Новые воз можности изучения стабильного пула углерода в длительных опытах с чистыми парами // Агрохи мия. 2011. № 12. С. 28–36.
2.Димо В.Н. Тепловой режим почв СССР. М.: Колос, 1972. 360 с.
3.Когут Б.М. Принципы и методы оценки содержа ния трансформируемого органического вещества в пахотных почвах // Почвоведение. 2003. № 3. С. 308–316.
4.Когут Б.М., Фрид А.С., Масютенко Н.П., Куваева Ю.В., Романенков В.А., Лазарев В.И., Холодов В.А. Дина мика содержания органического углерода в типич ном черноземе в условиях длительного полевого опыта // Агрохимия. 2011. № 12. С. 37–44.
5.Кудеяров В.Н., Демкин В.А., Гиличинский Д.А., Горячкин С.В., Рожков В.А. Глобальные изменения климата и почвенный покров // Почвоведение. 2009. № 9. С. 1027–1042.
6.Курганова И.Н., Лопес де Гереню В.О., Мякшина Т.Н.,
Сапронов Д.В., Кудеяров В.Н. Эмиссия СО2 из почв различных экосистем южно таежной зоны: анализ данных непрерывных 12 летних наблюдений // Доклады Академии наук АН. 2011. Т. 436. С. 843– 846.
7.Ларионова А.А., Золотарева А.Н., Евдокимов И.В., Быховец С.С., Кузяков Я., Бюггер Ф. Идентифика ция лабильного и устойчивого пулов органическо го вещества в агросерой почве // Почвоведение. 2011. № 6. С. 685–698.
8.Ларионова А.А., Стулин А.Ф., Занина О.Г.,
Евдокимов И.В., Хохлова О.С., Бюггер Ф., Шлотер М., Кудеяров В.Н. Распределение стабиль ных изотопов углерода в агрочерноземе при смене растительности с С3 типом фотосинтеза на моно культуру кукурузы // Почвоведение. 2012. № 8.
С.863–874.
9.Моргун Е.Г., Ковда И.В., Рысков Я.Г., Олейник С.А.
Возможности и проблемы использования методов геохимии стабильных изотопов углерода в почвен ных исследованиях // Почвоведение. 2008. № 3.
С.299–310.
10.Погода в Воронеже. http://www.pogoda.ru.net/climate/ 34123.htm
11.Семенов В.М., Иванникова Л.А., Кузнецова Т.В., Се менова Н.А., Тулина А.С. Минерализуемость орга нического вещества и углерод секвестрирующая емкость почв зонального ряда // Почвоведение. 2008. № 7. С. 819–832.
12.Blagodatskaya E., Kuzyakov Y. Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: critical review // Biology & Fertility of Soils. 2008. V. 45.
P.115–131.
13.Conen F., Leifeld J., Seth B., Alewell C. Warming miner alises young and old soil carbon equally // Biogeo sciences. 2006. V. 3. P. 515–519.
14.Davidson E.A., Janssens I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change // Nature. 2006. V. 440. P. 165–173. doi: 10.1038/nature04514
15.Fang C., Smith P., Moncrieff J., Smith J.U. Similar re sponse of labile and resistant soil organic matter pools to changes in temperature // Nature. 2005. V. 433.
P.57–59.
16.Hobie E.A., Werner R.A. Intramolecular, compound specific, and bulk carbon isotope patterns in C3 and C4 plants: a review and synthesis // New Phytol. 2004.
V.161. P. 371–385.
17.Karhu K., Fritze H., Tuomi M., Vanhala P., Spetz P., Kitunen V., Liski J. Temperature sensitivity of organic matter decomposition in two boreal soil profiles // Soil Biol. Biochem. 2010. V. 42. P. 72–82.
18.Kirschbaum M.U.F. The temperature dependence of or ganic matter decomposition – still a topic of debate // Soil Biol. Biochem. 2006. V. 38. P. 2510–2518.
19.Knorr W., Prentice I.C., House J.I., Holland E.A. Long term sensitivity of soil carbon turnover to warming // Nature. 2005. V. 433. P. 298–301.
20.Leifeld J., Fuhrer J. The temperature response of CO2 production from bulk soils and soil fractions is related to soil organic matter quality // Biogeochemistry. 2005.
V.75. P. 433–453.
21.Šantru° c ková H., Bird M.I., Lloyd J. Microbial process es and carbon isotope fractionation in tropical and temperate grassland soils // Functional Ecology. 2000.
V.14. P. 108–114.
22.Taylor A.F.S. Missing links – δ13C anomalies between substrates and consumers // New Phytol. 2008. V. 177.
P.845–847.
23.Tuomi M., Vanhala P., Karhu K., Fritze H., Liski J. Het erotrophic soil respiration – Comparison of different models describing its temperature dependence // Ecol. Modelling. 2008. V. 211. P. 182–190.
24.Vanhala P., Karhu K., Tuomi M., Sonninen E., Junger H., Fritze H., Liski J. Old soil carbon is more temperature sensitive than the young in the agricultural field // Soil Biol. Biochem. 2007. V. 39. P. 2967–2970.
25.Waldrop M.P., Firestone M.K. Altered utilization pat terns of young and old soil C by microorganisms caused by temperature shifts and N additions // Biogeochem istry. 2004. V. 67. P. 235–248.
ПОЧВОВЕДЕНИЕ № 7 2013

ПОЧВОВЕДЕНИЕ, 2013, № 7, с. 813–821
ФИЗИКА
ПОЧВ
УДК 631.437.213
БИОЭЛЕКТРИЧЕСКИЕ ПОТЕНЦИАЛЫ В СИСТЕМЕ ПОЧВА–РАСТЕНИЕ
© 2013 г. А. И. Поздняков
Факультет почвоведения МГУ им. М.В. Ломоносова, 119991, Москва, Ленинские горы
Поступила в редакцию 23.08.2011 г.
Проведено детальное изучение потенциалов в системе почва–растение относительно почв. Уста$ новлена зависимость величин потенциалов от вида растения и свойств почв. Дана первичная теоре$ тическая трактовка полученных данных.
Все растения, независимо от вида и состояния, всегда имеют отрицательный электрический потен$ циал по сравнению с почвой. Электрический потенциал травянистых растений в значительной мере зависит от площади поверхности листа. У таких растений, как лопух$репейник (Arctium lappa) и бор$ щевик сосновского (Heracléum sosnówskyi), наблюдаются большие по абсолютной величине отрица$ тельные значения электрических потенциалов – больше 100 мВ. Для всех растений наблюдается четкая дифференциация электрического потенциала – у цветка меньше, чем у листа, а у листа чаще всего меньше, чем у стебля. У таких растений, как подорожник (Plantago major), лопух (Arctium lap% pa), одуванчик (Taráxacum officinále) – в розетке листьев электрический потенциал выше, чем у са$ мого листа. Обнаруженные электрические потенциалы подчиняются сезонной динамике. Чем больше влажность почвы, тем меньше потенциал; только в случае одуванчика (Taráxacum officinаle) наблюдалась обратная зависимость. Можно полагать, что электрический потенциал между почвой и растением характеризует энергию жизнедеятельности растения.
Ключевые слова: почва, растение, электрофизические свойства, электрические потенциалы.
DOI: 10.7868/S0032180X13070095
ВВЕДЕНИЕ
Одним из главных абиотических факторов окружающей природной среды являются физиче$ ские поля, заполняющие среду обитания и воздей$ ствующие на живую и неживую природу. Воздей$ ствие физических полей определяет большинство процессов, протекающих в биосфере. Биосферно$ ландшафтные взаимоотношения всегда сопро$ вождаются электрическими явлениями и генери$ рованием электрических полей.
Важным аспектом этой проблемы является взаимодействие почв с электромагнитными по$ лями атмосферы, влияние на которые оказывают электрические свойства почв, их способность к поляризации, то есть к неравномерному распре$ делению электрических потенциалов в биосфере, ландшафте, почве и растении.
Что касается самих электрических полей в почвах, то они изучены уже достаточно подробно, в том числе и нами [7–12]. Выяснено что прояв$ ление естественного электричества в почвах весь$ ма разнообразно: это диффузионно$адсорбцион$ ные, фильтрационные, электрохимические и другие поля. Исходя из распространенности и продолжительности действия, по$видимому, можно считать, что основной составляющей есте$ ственных электрических полей (потенциалов) в
почвах являются диффузионно$адсорбционные электрические поля, возникающие вследствие перераспределения ионов под действием почво$ образовательных процессов [10–12]. Эти поля обусловлены диффузией и адсорбцией ионов, ко$ торые в свою очередь в значительной мере зави$ сят от содержания воды в почве и количества ионогенных соединений.
Достаточно хорошо известны биофизические работы по электрофизиологии самих растений. Работ по электрическим потенциалам на перехо$ де почва–растение нам обнаружить не удалось ни в отечественной, ни в зарубежной литературе. Удалось найти лишь работы по поляризации внутри самого растения [2, 16], но не об электри$ ческих потенциалах на переходе почва–растение. Так, например, Маслоброд пишет, что электро$ физиологическая или биоэлектрическая поляр$ ность растений наглядно проявляется при иссле$ довании топографии биоэлектрических потенци$ алов, а также при регистрации электрических ответов растения на раздражение [4].
В первом случае объект находится в стацио$ нарном невозбужденном состоянии, и его биоло$ гическая полярность выражается в различии ме$ таболических потенциалов или потенциалов по$ коя между отдельными органами, тканями,
813

814 |
ПОЗДНЯКОВ |
клетками. Заметим, что эти различия потенциа$ лов изучены только внутри растения, а не по от$ ношению к почве, когда большую роль играют свойства и особенности самих почв. Метаболиче$ ские потенциалы самих растений, несут полез$ ную информацию о растении как о целостной по$ ляризованной системе и объективно характеризу$ ют ее жизнеспособность. Но макроэлектрические потенциалы всего растения по отношению к поч$ ве более информативны, так как характеризуют поведение объединенных сред и их функциони$ рование, что отражает электрическое состояние всего растения в сравнении с почвой, в том числе и внутренних тканей, заряд которых противопо$ ложен наружным. Топография же метаболиче$ ских потенциалов в самом растении характеризу$ ет лишь определенное морфологическое состоя$ ние только растения, а также динамику этого состояния, то есть определенный этап онтогене$ за. Измеряя макропотенциал поверхности любой части растительного объекта по сравнению с по$ тенциалом почвы, получаем в первую очередь ин$ формацию о потенциале покоя группы клеток, составляющих эту поверхность и в какой$то мере потенциал их динамики.
Поэтому можно ожидать скачка потенциалов в системе почва–растение, который будет свиде$ тельствовать одновременно о целой гамме взаи$ модействующих биоэлектрических потенциалов, протекающих в почве и растении. В результате всевозможных наложений и корреляций ритмик макробиоэлектрических потенциалов возникают относительно стабильные по топографии зоны с различными уровнями биоэлектрической актив$ ности. Это находит свое отражение в наличии макробиоэлектрической полярности растения по отношению к почве. Отдельные типы биоэлек$ трической полярности, по$видимому, несут раз$ личную информацию о растительном организме и почве. Макробиологическая полярность отра$ жает присущее всякому живому организму состо$ яние постоянного возбуждения или “устойчивого неравновесия” по отношению к среде обитания, одной из составляющих экосистемы – почве. Это возбуждение имеет форму срочного или медлен$ ного изменения стационарного уровня биопотен$ циалов с различными оттенками перехода одного состояния в другое. Это тоже должно служить ос$ нованием для возникновения потенциалов между почвой и растением.
Первые измерения указали на существование таких электрических потенциалов между почвой и растением [24]. В то же время оставались нере$ шенными многие вопросы, основными из кото$ рых являются следующие: 1 – насколько повсе$ местно распространены эти электрические явле$ ния, 2 – проявляются ли они на всех почвах и для всех растений, 3 – насколько они стабильны и ди$ намичны, 4 – от каких факторов они зависят? Ре$
шению этих вопросов и посвящена настоящая ра$ бота.
ОБЪЕКТЫ И МЕТОДЫ
Большинство исследований проводились в окрестностях Приокско$Террасного биосферного заповедникa в Серпуховском р$не Московской обл., Татарской лесной опытной станции (ТатЛОС) и на сельскохозяйственных торфяных ландшафтах Дмитровского филиала ВНИИМЗ Яхромской поймы Дмитровского р$на Московской обл. (Россия), а также в условиях острова Минданао
1
близ г. Замбоанга (Филиппины) .
При измерениях особое внимание было уделе$ но оценке электрических явлений, процессов и полей, формирующихся при почвообразовании,
иизучению воздействия их в системе почва–рас$ тение при многодневной и суточной динамике; выявлению электрических барьеров на границе почва–растение, их взаимосвязи с электрохими$ ческими характеристиками и электрополяриза$ цией почвы. Были изучены параметры, направ$ ленность и поведение электрических полей рас$ тений по отношению к верхним горизонтам дерновых и дерново$подзолистых почв основных естественных и искусственных экосистем этих регионов в реальных условиях. Измерены элек$ трические потенциалы этого перехода более чем 80 видов растений в естественных биогеоценозах
икультурных растений (в основном овощных) в агроценозах на почвах основных генетических типов Нечерноземья.
Были изучены также потенциалы перехода почва–растение некоторых тропических расте$ ний в реальных условиях на Филлипинах (остров Минданао).
Измерения электрических потенциалов пере$ хода почва–растение проводили по ранее разра$ ботанной методике для измерения потенциалов почв и растений, используя неполяризующиеся электроды [6–8, 13, 19] и руководствуясь рядом рекомендаций по электродам приведенных в за$ рубежных работах [20–22].
Измерения осуществляли методом отведения потенциала с помощью двух неполяризующихся хлор$серебряных неполяризующихся электродов с насадками из загущенного агарового раствора 1% хлорида калия [13, 19]. Один электрод уста$ навливали в почве, в непосредственной близости от растения, а вторым в процессе измерения при$ касались к стволу (стеблю) растения или его ча$ стям. Прибор, с помощью которого регистриро$ вали потенциалы, – милливольтметр LandMapper ERM$02 [WWW.Landviser.net] с входным (внут$ ренним) сопротивлением 5 МОм. Перед каждой серией измерений проводили оценку поляриза$
1Ряд измерений получен совместно с А.С. Пуряевым, М.П. Микус и Н.И. Мирсияповым.
ПОЧВОВЕДЕНИЕ № 7 2013
БИОЭЛЕКТРИЧЕСКИЕ ПОТЕНЦИАЛЫ В СИСТЕМЕ ПОЧВА–РАСТЕНИЕ |
815 |
ции самих электродов [13, 19, 20–22], путем за$ земления обоих электродов в почве рядом друг с другом (табл. 1). Поляризация электродов учитыва$ лась при дальнейшей обработке данных и их кор$ ректировке путем алгебраического сложения непо$ средственно измеренных потенциалов в системе почва–растение и поляризации электродов.
Заметим также, что хотя во всех случаях изме$ ряли разности электрических потенциалов, тем не менее, при описанной выше схеме измерений, можно судить о том, потенциал какого объекта – почвы или растения – больше или меньше, по$ скольку измерить абсолютную величину электри$ ческого потенциала не представляется возмож$ ным в принципе.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В результате первичного анализа полученных данных было установлено, что независимо от ви$ да растений (травянистые, древесные, кустарни$ ковые) и вида почвы, на которой они произраста$ ют, а также каких бы то ни было других причин, электрический потенциал между почвой и расте$ нием всегда отрицательный (табл. 1–4).
Э л е к т р и ч е с к и е п о т е н ц и а л ы м е ж $ д у т р а в я н и с т ы м и р а с т е н и я м и и п о ч в о й. Наиболее обширной группой растений, у которых проводили измерения электрических по$ тенциалов, были травянистые растения (табл. 1, 3).
Обнаруженные отрицательные разности элек$ трических потенциалов между почвой и травяни$ стыми растениями подчиняются сезонной дина$ мике (рис. 1, табл. 2). Весной электрические по$ тенциалы между почвой и растением более разительны, то есть значительно больше по абсо$ лютной величине, но тоже отрицательные. Летом эти различия уменьшаются и сильно колеблются во время измерений. Еще больше они уменьша$ ются к осени, когда становятся самыми низкими, то есть потенциалы почвы и растения выравнива$ ются и стабилизируются. Подобная общая дина$ мика, по$видимому, связана со стадиями разви$ тия растения, а не с погодными условиями. Так как электрические потенциалы, измеренные у молодых растений летом, оказались близкими по значениям к потенциалам, измеренным в мае (табл. 2), когда погодные условия разные.
Установлено, что при повышенной влажности почвы, например, после дождя, электрический потенциал меньше по абсолютной величине, чем потенциал, измеренный при других более сухих условиях. Между влажностью и потенциалом об$ наружена хорошая прямая корреляция и только в случае исследований одуванчика (Taraxacum offi% cinale) наблюдается обратная зависимость.
У всех растений обнаружена четкая дифферен$ циация – электрический потенциал по абсолют$ ной величине у цветка меньше, чем у листа, а у листа чаще всего меньше, чем у стебля (табл. 1).
|
|
|
Клевер |
Подорожник |
|||
|
V |
|
VII |
IX |
V |
VII IX |
|
|
|
VI |
VIII |
VI |
VIII Месяц |
||
|
0 |
|
|
|
|
0 |
|
|
–20 |
|
|
|
|
–40 |
|
|
–40 |
|
|
|
|
–80 |
|
|
|
|
|
|
|
|
|
|
–60 |
|
|
|
|
–120 |
|
|
|
|
|
|
|
|
|
|
|
|
Осот |
|
Одуванчик |
||
|
V |
VII |
IX |
V |
VII |
||
мВ |
VI |
VIII |
X |
VI |
VIII |
||
0 |
|
|
|
|
0 |
|
|
потенциал, |
|
|
|
|
|
||
–20 |
|
|
|
|
–20 |
|
|
|
–40 |
|
|
|
|
–40 |
|
|
|
|
|
|
–60 |
|
|
|
–60 |
|
|
|
|
|
|
|
|
|
|
|
–80 |
|
|
|
–80 |
|
|
|
|
|
|
Электрический |
|
Пижма |
Полынь (лист) |
||||
|
|
||||||
V |
VII |
IX |
|
V |
VII |
||
|
VI |
VIII |
X |
VI |
VIII |
||
|
0 |
|
|
|
|
0 |
|
|
–20 |
|
|
|
|
–20 |
|
|
|
|
|
|
–40 |
|
|
|
|
|
|
|
|
|
|
|
–40 |
|
|
|
|
–60 |
|
|
–60 |
|
|
|
|
–80 |
|
|
|
|
|
|
|
|
|
|
Тысячелистник |
|
|
||||
|
V |
VII |
IX |
|
|
|
|
|
VI |
VIII |
X |
|
|
||
|
0 |
|
|
|
|
|
|
|
–20 |
|
|
|
|
|
|
–40  –60
–60 

Сезонная динамика электрических потенциалов в си$ стеме почва–растение: клевера (Trifolium repens), по$ дорожника (Plantago major), осота (Sonchus arvensis), одуванчика (Taraxacum officinаle), пижмы (Tanacetum balsamita), полыни (Artemisia absinthium), тысячелист$ ника (Achillea millefolium).
Например, у цветка клевера электрический по$ тенциал составляет –30 мВ, у листа – –39 мВ, а у стебля –44 мВ. У таких растений, как подорож$ ник (Plantago major), лопух (Arctium lappa), оду$ ванчик (Taraxacum officinale), в розетке листьев разность потенциалов больше, чем у отдельного листа. Электрические различия в потенциалах травянистых растений в сильной мере зависят и от площади поверхности листа. У таких растений, как лопух (Arctium lappa) и борщевик сосновского (Heracléum sosnówskyi), наблюдаются очень большие по абсолютной величине значения электрических потенциалов: больше минус 100 мВ.
У культурных растений наблюдалось следую$ щее распределение макроэлектропотенциалов – у плода самый низкий, больше у листьев, и самый большой потенциал у стебля или корнеплода (табл. 1).
ПОЧВОВЕДЕНИЕ № 7 2013