Материал: 1010-pochvovedenie-2013-7

816 |
ПОЗДНЯКОВ |
|
||
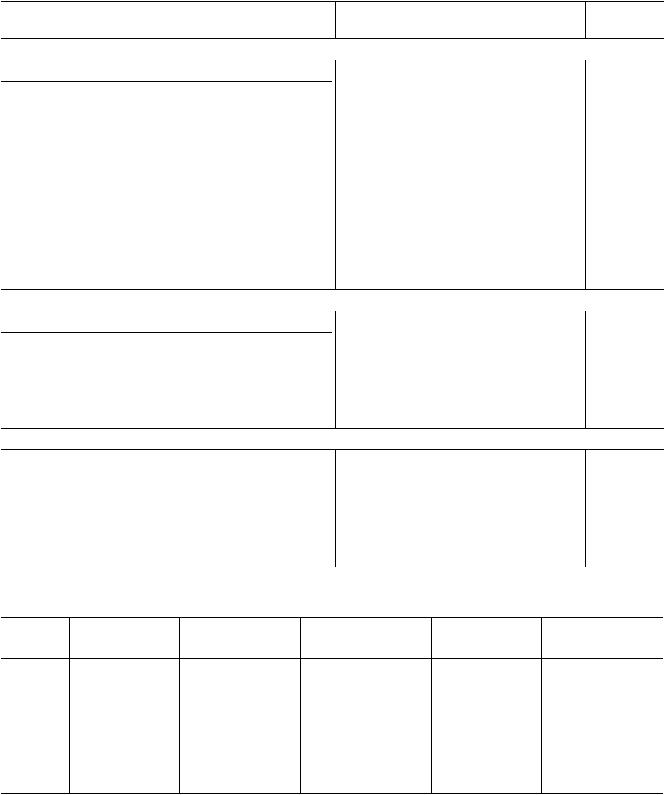
Таблица 1. Естественные электрические потенциалы между почвой и растением |
|
|||
Растение |
Потенциал, |
Растение |
Потенциал, |
|
мВ |
мВ |
|||
|
|
|||
Пойма р. Яхромы, 8 кв. стационара “Дальний” ВНИИМЗ |
|
|||
Поляризация электродов |
–0.56 |
Поляризация электродов |
–4.6 |
|
Калужница Caltha palustris |
–182 |
Клевер Trifolium repens |
–49 |
|
Ива (побег) Salix |
–50 |
Одуванчик Taraxacum officinаle (цветок) |
–68 |
|
Ива (почка) Salix |
–17 |
Осот Sоnchus arensis |
–40 |
|
|
|
Пижма Tanacetum balsamita |
–21 |
|
Приокско$Террасный заповедник |
|
|||
На поле |
|
В сосняке |
|
|
Поляризация электродов |
–1.4 |
Подорожник Plantago major |
–92 |
|
Малина Rubus vulgatus |
–70 |
Манжетка Alchemilla vulgaris |
–107 |
|
Одуванчик Taraxacum officinаle |
–50 |
Одуванчик Taraxacum officinаle |
–80 |
|
Осот Sоnchus arensis |
–67 |
Сосна (50 лет ) Pinus sylvestris |
–10 |
|
Осот Sоnchus arensis |
–80 |
Одуванчик (розетка) Taraxacum officinаle |
–50 |
|
Одуванчик (розетка) Taraxacum officinаle |
–80 |
Осот Sоnchus arensis |
–96 |
|
Сосна (15 лет, ствол) Pinus sylvestris |
–10 |
Одуванчик (бутон) Taraxacum officinаle |
–45 |
|
Сосна (ствол) Pinus sylvestris |
–1.9 |
Чистяк (листок) Ranunculus ficaria |
–60 |
|
Сосна (новый побег) Pinus sylvestris |
–19 |
Чистяк (цветок) Ranunculus ficaria |
–13 |
|
Береза (2 года) Betula pendula |
–30 |
Одуванчик (цветок) Taraxacum officinаle |
–30 |
|
Клевер Trifolium repens |
–68 |
Манжетка (лист) Alchemilla vulgaris |
–68 |
|
Одуванчик (цветок) Taraxacum officinаle |
–19 |
Ольха (апек) Alnus glutinosa |
–37 |
|
Крестоцветые (цветок) |
–32 |
Ольха (листья) Alnus glutinosa |
–60 |
|
Живучка Ajuga reptans |
–69 |
|
|
|
|
У шоссе, 20.07.08 г. |
|
||
Поляризация электродов |
–4.64 |
Клевер Trifolium repens |
–26.2 |
|
Одуванчик (розетка) Taraxacum officinаle |
–90.5 |
Одуванчик (бутон) Taraxacum officinаle |
–53.2 |
|
Одуванчик (цветок) Taraxacum officinаle |
–74.1 |
Одуванчик (шарик) Taraxacum officinаle |
–23.7 |
|
Пижма Tanacetum balsamita |
–81.5 |
Вероника Veronica officinalis |
–51.6 |
|
Береза (Н = 2.3 м) Betula pendula |
–48.7 |
Манжетка Alchemilla vulgaris |
–45.1 |
|
Тысячелистник Achillea millefolium |
–59.9 |
Вероника (цветок) Veronica officinalis |
–3.01 |
|
|
У шоссе, 27.07.08 г. |
|
||
Поляризация электродов |
3.3 |
Яснотка (цветок) Lamium album |
–40 |
|
Донник (стебель) Melilotus albus |
–19.8 |
Яснотка (стебель) Lamium album |
–65.7 |
|
Клевер луговой (цветок) Trifolium pratense |
–16.4 |
Герань (лист) Geranium sylvaticum |
–77.1 |
|
Клевер луговой (лист) Trifolium pratense |
–20.8 |
Герань (цветок) Geranium sylvaticum |
–47 |
|
Клевер луговой (стебель) Trifolium pratense |
–18.5 |
Вероника (цветок) Veronica officinalis |
–16.2 |
|
Жерушник Rorippa amphibia |
–46.5 |
Клевер ползучий (цветок) Trifolium repens |
–9.38 |
|
Ежа Dactylis glomerata |
–10.5 |
Мятлик Poa pratensis |
–11.7 |
|
Манжетка Alchemilla vulgaris |
–44.7 |
Ромашка пахучая Matricaria discoidea |
– |
|
Цикорий (лист) Cichorium intybus |
–43.1 |
Тысячелистник (стебель) Achillea millefolium |
–36.8 |
|
Подорожник Plantago major |
–62.7 |
Лопух (лист) Arctium lappa |
–67.7 |
|
Лисохвост Alopecurus pratensis |
–16.4 |
Лопух (розетка) Arctium lappa |
–123 |
|
Одуванчик (лист) Taraxacum officinаle |
–35.6 |
Хвощ Equisetum fluviatile |
–7.8 |
|
Купырь (лист) Anthriscus sylvestris |
–35.5 |
Короставник (цветок) Scabiosa arvensis |
–65.7 |
|
Купырь (цветок) Anthriscus sylvestris |
–24.8 |
Короставник (стебель) Scabiosa arvensis |
–48.7 |
|
Щавель Rumex confertus |
–48.9 |
Донник (цветок) Melilotus albus |
–6.7 |
|
ПОЧВОВЕДЕНИЕ № 7 2013
БИОЭЛЕКТРИЧЕСКИЕ ПОТЕНЦИАЛЫ В СИСТЕМЕ ПОЧВА–РАСТЕНИЕ |
817 |
|||
Таблица 1. Окончание |
|
|
|
|
Растение |
Потенциал, |
Растение |
Потенциал, |
|
мВ |
мВ |
|||
|
|
|||
|
У шоссе, 10.08.04 г. |
|
||
Поляризация электродов |
–13.2 |
Осот (лист) Sоnchus arensis |
–11.9 |
|
Тысячелистник (лист) Achillea millefolium |
–12.1 |
Осот (стебель) Sоnchus arensis |
–14.9 |
|
Подорожник (лист) Plantago major |
–25 |
Бодяк полевой (цветок) Cirsium arvense |
–25.2 |
|
Марьянник (цветок) Melampyrum pratense |
–9.8 |
Бодяк полевой (стебель) Cirsium arvense |
–39.7 |
|
Марьянник (стебель) Melampyrum pratense |
–13.3 |
Пижма (стебель) Tanacetum balsamita |
–32.9 |
|
Горошек (лист) Vicia cracca |
–14.9 |
Пижма (лист) Tanacetum balsamita |
–23.5 |
|
Горошек (стебель) Vicia cracca |
–20.7 |
Пижма (цветок) Tanacetum balsamita |
–9.2 |
|
Клевер ползучий (лист) Trifolium repens |
–9 |
Одуванчик (розетка) Taraxacum officinаle |
–40 |
|
Клевер ползучий (цветок) Trifolium repens |
–6.06 |
Одуванчик (лист) Taraxacum officinаle |
–67.4 |
|
Клевер луговой (лист) Trifolium pratense |
–10.8 |
Чистец болотный (цветок) Stachys palustris |
–7 |
|
Полынь (лист) Artemisia absinthium |
–13.3 |
Чистец болотный (стебель) Stachys palustris |
–18 |
|
Полынь (стебель) Artemisia absinthium |
–30.7 |
Чистец болотный (лист) Stachys palustris |
–37.6 |
|
|
У шоссе, 20.08.04 г. |
|
||
Поляризация электродов |
2.8 |
Полынь (стебель) Artemisia absinthium |
–38 |
|
Полынь (лист) Artemisia absinthium |
–23 |
Клевер луговой (лист) Trifolium pratense |
–5 |
|
Подорожник (лист) Plantago major |
–47 |
Клевер луговой (стебель) Trifolium pratense |
–9 |
|
Одуванчик (лист) Taraxacum officinаle |
–51 |
Горошек (лист) Vicia cracca |
–12 |
|
Осот (стебель) Sоnchus arensis |
–42 |
Щавель (лист) Rumex confertus |
–47.3 |
|
Осот (лист) Sоnchus arensis |
–19 |
Пижма (лист) Tanacetum balsamita |
–34 |
|
|
У шоссе, 12.09.04 г. |
|
||
Поляризация электродов |
–7.8 |
Полынь (стебель) Artemisia absinthium |
–4.3 |
|
Подорожник (лист) Plantago major |
–6.7 |
Клевер луговой (лист) Trifolium pratense |
–5.6 |
|
Одуванчик (лист) Taraxacum officinаle |
–12.5 |
Тысячелелистник (лист) Achillea millefolium |
–11 |
|
Осот (стебель) Sоnchus arensis |
–26.3 |
Щавель (стебель) Rumex confertus |
–8.9 |
|
Осот (лист) Sоnchus arensis |
–16.4 |
Щавель (лист) Rumex confertus |
–6.3 |
|
Полынь (лист) Artemisia absinthium |
–4.2 |
Пижма (лист) Tanacetum balsamita |
–13.4 |
|
Таблица 2. Средние значения электрических потенциалов между почвой и растением (мВ) по месяцам вегетации
Месяц |
Подорожник |
Пижма |
Одуванчик |
Клевер |
Тысячелистник |
|
(Plantago major) |
(Asteraceae balsamita) (Taraxacum officinаle) |
(Trifolium repens) |
(Achillea millefolium) |
|||
|
||||||
Май |
–92.0 |
–51.3 |
–80.0 |
–65.2 |
–60.0 |
|
Июнь |
–98.0 |
–33.0 |
–65.0 |
–43.3 |
–35.7 |
|
Июль |
–51.4 |
–26.5 |
–61.6 |
–29.2 |
–21.4 |
|
Август |
–44.1 |
–37.9 |
–51.9 |
–13.6 |
–16.1 |
|
Сентябрь |
–28.3 |
–17.1 |
–24.8 |
–4.8 |
–15.0 |
|
Октябрь |
–13.0 |
–12.4 |
–5.1 |
–10.8 |
–7.4 |
|
Э л е к т р и ч е с к и е п о т е н ц и а л ы м е ж $ д у п о ч в о й и д р е в е с н ы м и р а с т е н и я $ м и. По величинам разностей электрических по$
тенциалов между почвой и растением (стволом) древесные породы принципиально не отличают$ ся от травянистых. В одном и том же биогеоцено$
4 ПОЧВОВЕДЕНИЕ № 7 2013
818 |
ПОЗДНЯКОВ |
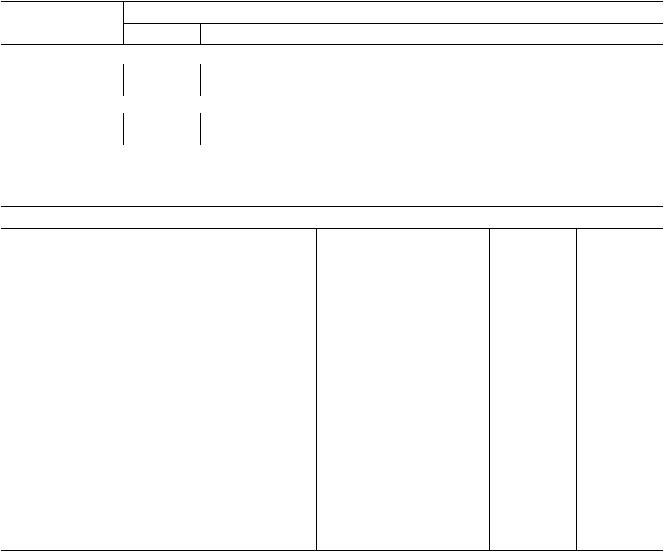
Таблица 3. Динамика разности электрических потенциалов (мВ) между торфяной почвой и корнеплодами мор$ кови Нантской в августе 2008 г. на восьмом квартале Дмитровского филиала ВНИИМЗ (среднее значение из пя$ ти повторностей)
Размер корнеплода |
|
|
Время наблюдения в минутах |
|
|
||
|
|
|
|
|
|
|
|
моркови, см |
0 |
10 |
20 |
30 |
45 |
60 |
90 |
|
|||||||
|
|
Неповрежденные растения, август 2008 г. |
|
|
|
||
15–20 |
–127 |
–120 |
–101 |
–113 |
–100 |
–112 |
–122 |
20–25 |
–150 |
–144 |
–157 |
–148 |
–156 |
–147 |
–132 |
|
Поврежденные надрезами корнеплоды, июль 2008 г. |
|
|
||||
10–18 |
–48 |
–30 |
–38 |
–42 |
–37 |
–41 |
|
20–25 |
–110 |
–94 |
–77 |
–76 |
–65 |
–67 |
|
Таблица 4. Естественный электрический потенциал (мВ) между почвой и некоторыми тропическими растения$ ми Филиппин (среднее из 5–7 повторностей). Здесь и в табл. 5, 6 приведены средние значения потенциалов для отдельных растений
Вид растения |
|
Разность потенциалов, мВ |
|
||
Мимоза стыдливая (Mimosa pudica) |
–278 |
–182 |
–149 |
–258 |
–312 |
Растения бананов (Músa): |
–72 |
–58 |
–49 |
–58 |
–86 |
свежесрубленные растения |
–34 |
–28 |
–22 |
–24 |
|
стебель бананового растения |
–52 |
–48 |
–56 |
|
|
Папайя (Carica papaya, подрост) |
–87 |
–92 |
–82 |
–78 |
|
Дерево какао (Theobroma cacao) |
–27 |
–32 |
–29 |
|
|
Хлебное дерево (Artocarpus altilis): |
–128 |
–132 |
–135 |
|
|
через 15 мин после засечки |
–88 |
–92 |
–88 |
|
|
листья |
–42 |
–46 |
–48 |
–52 |
|
Кокосовая пальма (Cocos nucifera): |
–89 |
–79 |
–91 |
|
|
через 15 мин после надреза |
–89 |
–92 |
–90 |
|
|
Бананы (Músa): |
|
|
|
|
|
через 2 дня после повреждения: |
–57 |
–58 |
–62 |
|
|
поврежденные растения |
–34 |
–38 |
–42 |
–46 |
|
внутри поврежденной пальмы |
–56 |
–59 |
–52 |
|
|
наружные стеблевые листья |
–28 |
–33 |
–34 |
|
|
Ананас (Ananas, листья): |
–24 |
–28 |
–22 |
|
|
шипы на листьях ананаса |
–32 |
–36 |
–29 |
|
|
плоды (срез) |
–52 |
–58 |
–46 |
–48 |
|
зе древесные растения могут как превосходить часть травянистых растений по этому показате$ лю, так и уступать им (табл. 1, 4–6). Так, напри$ мер, у осины (Pópulus trémula) потенциал достига$ ет –120 мВ, а у сосны (Pinus sylvestris) составляет всего минус 10 мВ (табл. 4, 5). Кроме того, в отли$ чие от трав, не обнаружено различий величин по$ тенциала между деревьями разного возраста. Од$ нако среди разновозрастных частей одного дерева различия носят тот же характер – более молодые ветви имеют большие абсолютные значения по$ тенциала, чем ствол и старые ветви (табл. 5).
Электрические потенциалы системы почва– растения находятся в очень хорошей зависимо$ сти от категории состояния древесных пород (табл. 6). Так, для липы (Tilia europaea) первой (лучшей) категории величина потенциала со$ ставляет около –30…–50 мВ, для третьей катего$
рии – не больше –20…–30 мВ, а для четвертой (самой плохой) падает до 3–10 мВ. Интересно, что соответствующие потенциалы сохраняются и в пне от свежесрубленного дерева, у которого не$ которое время продолжается сокодвижение.
На данном этапе исследований мы пришли к выводу, что электрический потенциал между поч$ вой и растением в какой$то мере характеризует не только энергию состояния и жизнедеятельности растения, но и потребности растения в воде в конкретный момент времени.
Наблюдения за комнатными растениями по$ казали, что когда растению требуется полив, оно показывает высокий по абсолютной величине от$ рицательный потенциал по отношению к почве. После полива сначала наблюдается резкое увели$ чение разности потенциалов по абсолютной ве$
ПОЧВОВЕДЕНИЕ № 7 2013

БИОЭЛЕКТРИЧЕСКИЕ ПОТЕНЦИАЛЫ В СИСТЕМЕ ПОЧВА–РАСТЕНИЕ |
819 |
Таблица 5. Электрические потенциалы (мВ) между гумусовым горизонтом дерново$подзолистых почв и некото$ рыми породами деревьев, слагающими БГЦ (каждое значение потенциала – среднее из семи повторностей)
Порода дерева |
|
|
Разность потенциалов на высоте 1.5 м |
|
|
||
Рябина (Sorbus) |
–75 |
–78 |
–65 |
–69 |
–82 |
–67 |
–69 |
Осина (Pópulus trémula) |
–100 |
–120 |
–114 |
–117 |
–123 |
–117 |
–110 |
личине, вероятно, показывающее, что растение всасывает воду. Потом происходит уменьшение потенциала до величины меньшей по абсолютной величине, чем он был до полива. Для каждого растения характерно свое время, когда потенциал после полива увеличивается.
Как видим, для каждого вида растений суще$ ствует определенная величина электрического потенциала, зависящая от физиологического со$ стояния этого растения и от условий окружающей среды, в первую очередь почвы. По нашему мне$ нию, первичный потенциал является характери$ стикой не только роста и развития растения, но и водообеспеченности. Если электрический потен$ циал связан с влагообеспеченностью, то он свя$ зан и с транспирацией, поскольку, как показано в работе Судницына [14], влагообеспеченность и транспирация тесно взаимосвязаны.
Для того, чтобы пояснить причины такого рас$ пределения электрических явлений и потенциа$ лов в системе почва–растение, привлечем совре$ менные представления из смежных областей зна$ ния [15]. Обычно внутри однородной среды положительные и отрицательные заряды в доста$ точно большом однородном объеме одинаковы, то есть суммарный заряд равен нулю. Однако, ес$ ли по каким$либо причинам внутри системы рас$ пределение зарядов неоднородно (в одной обла$ сти больше “+”, в другой “–”), то между этими областями возникает разность потенциалов [15].
Простейшим механизмом такой электрохими$ ческой самоорганизации ионов в водной среде является образование ионных кластеров. Пред$ положим, например, что вокруг центрального по$ ложительного иона под действием сил электро$ статического притяжения оказались один или не$ сколько отрицательных противоионов. Такой кластер уже является источником разности по$ тенциалов, однако тепловое движение будет раз$ рушать структуру кластера, то есть он имеет опре$ деленное время жизни [23, 25]. Теория ионных кластеров была развита Дебаем и Гюккелем [8–10, 12]. Она получила признание в электрохимии и биофизике и может быть использована для объяс$ нения возникновения разности потенциалов на границе почва–растение и в самой почве.
Важным и достаточно значимым механизмом электрогенеза является образование широко из$ вестного двойного электрического слоя с фикси$ рованным электрическим зарядом на поверхно$ сти почвенной частицы или клеточной мембра$
ны. На внешней стороне таких образований формируется избыток положительных, а на внут$ ренней – избыток отрицательных зарядов при ну$ левом суммарном заряде объекта. Соответствен$ но, концентрация противоионов в водном рас$ творе вблизи раздела сред возрастает, формируя два слоя – ближайший к поверхности, состоящий из прочносвязанных ионов (слой Гельмгольца) и более удаленный диффузный слой (слой Чэпме$ на–Гуи) [15].
Теория диффузного двойного слоя аналогична теории Дебая–Гюккеля и обосновывает распреде$ ление электрических зарядов в двойном электри$ ческом слое на основании энергетического рас$ пределения частиц Максвелла–Больцмана [15].
Подобна и теория равновесного потенциала Нернста частично проницаемой для ионов био$ логической мембраны. Широко распространен$ ная формула Нернста позволяет объяснить и ко$ личественно описать образование потенциала покоя на мембранах клеток и клеточных орга$ нелл, а также процесс генерации и распростране$ ния потенциала действия [4, 5, 10].
В почвах роль такой мембраны играют грани$ цы раздела между горизонтами, например, элю$
Таблица 6. Зависимость электрических потенциалов (мВ) в системе почва–дерево для липы мелколистной (Tilia cordata) от разного состояния дерева (среднее из пяти повторностей)
Категория |
Разность потенциалов ствола дерева |
||
|
на высоте 1.5 м, мВ |
|
|
состояния |
cредние значения потенциалов |
||
дерева |
|||
|
|
отдельных деревьев |
|
1 |
–45 |
–50 |
–59 |
1 |
–44 |
–50 |
–33 |
1 |
–33 |
–36 |
–33 |
1 |
–52 |
–54 |
–44 |
1 |
–40 |
–45 |
–40 |
1 |
–44 |
–50 |
–31 |
2 |
–38 |
–42 |
–38 |
2 |
–42 |
–49 |
–40 |
3 |
–39 |
–34 |
–27 |
3 |
–26 |
–26 |
–25 |
3 |
–24 |
–34 |
–30 |
3 |
–27 |
–5 |
–19 |
4 |
–23 |
–17 |
–13 |
4 |
–10 |
–3 |
–18 |
ПОЧВОВЕДЕНИЕ № 7 2013 |
4* |
820 |
ПОЗДНЯКОВ |
виального–иллювиального, засоленного–неза$ соленного, оглеенного–оподзоленного и других подобных с резкими различиями диффузионно$ адсорбционных свойств [17, 19, 22, 23, 25].
Все вышеперечисленные процессы и явления в той или иной мере вносят свой вклад в форми$ рование разности потенциалов между почвой и растением на границе раздела почва–корень рас$ тения и далее для всего растения со всеми его со$ ставными частями.
На наш взгляд, рассмотренные разности по$ тенциалов в системе почва–растения являются яркими примерами обобщенной трактовки воз$ никновения разности потенциалов, сформулиро$ ванной великим физиком XX в. И.Е. Таммом. По поводу формирования электрических потенциа$ лов в самых различных средах он писал: … “в по$ граничном слое между любыми соприкасающи$ мися проводниками различного химического со$ става возникают разности потенциалов, величина которых зависит в первую очередь от химической природы соприкасающихся проводников…” [15]. Обычно частные проявления аналогичных разно$ стей электрических потенциалов, например, мембраны клеток корня, описывают особенно$ стями транспорта каких$либо одних ионов, на$ пример, калия через мембрану, роль других ионов, особенно натрия, игнорируется или при$ уменьшается, на что указывалось нами в преды$ дущих работах [5–8]. В ряде случаев это может быть и справедливо. Но при измерениях потенци$ алов между почвой и растением, когда не проис$ ходит непосредственного пересечения мембран$ ных формирований растения, оно не нарушается. Причина устойчивых разностей потенциалов кро$ ется в различиях состава ионогенных соединений в разных частях этой системы, то есть объясняется различиями концентраций этих соединений в поч$ ве и растении.
Согласно отрицательной разности потенциа$ лов между почвой и растением, в растениях таких ионогенных соединений больше, чем в почвах, поэтому природа этих потенциалов в системе почва–растение сродни диффузионно$сорбци$ онным потенциалам, наблюдаемым в самых раз$ личных средах, в том числе в почвах, самих расте$ ниях и, как видим, в объединяющей их системе почва–растение. Этот электрический потенциал может в свою очередь определять характер транс$ порта ионов в растение.
На данном этапе исследований представляет$ ся, что поверхностный электрический макробио$ потенциал растений на переходе почва–расте$ ние, по$видимому, отражает электрическое со$ стояние всего растения, в том числе ЭДС внутренних тканей, заряд которых противополо$ жен наружным тканям. Топография метаболиче$ ских потенциалов в растении отражает, с одной стороны, определенное морфологическое строе$ ние и состояние растения, с другой стороны – ди$
намику этого состояния, то есть определенный этап онтогенеза. Измеряя макробиопотенциал поверхности любой части растительного объекта, мы получаем информацию об обобщенном по$ тенциале – потенциале покоя группы клеток, со$ ставляющих эту поверхность, а также потенциале физиологических процессов, таких, как сокодви$ жение, которые очень сильно изменяют подвиж$ ность и количество электрических зарядов в раз$ ных частях системы. Особенно это изменение сказывается при переходах в системе почва–рас$ тение в разное по погодным условиям время.
Поэтому кроме потенциалов статического со$ стояния добавляется мобильная составляющая за счет движения в растении этих растворов, кото$ рая гасится при увлажнении почвы или при поли$ ве растения, а также осенью, когда интенсивность сокодвижения уменьшается и электрические по$ тенциалы в системе почва–растение становятся спокойней – мало флуктуируют и практически выравниваются.
Итак, электропизм системы почва–растение не$ сет информацию не только о состоянии растений и их отношении к почвам, но свидетельствует о физио$ логическом состоянии растений. Электропизм ва$ жен не только с теоретической, но и с практической точки зрения, например, его можно использовать, учитывая при электровоздействии на растения – электростимуляции [11, 16–19, 24].
Для того, чтобы активно использовать явление электропизма необходимо научиться правильно его трактовать. Тем не менее, ряд результатов по$ добных исследований находят свое применение уже сейчас [3, 17]. В первую очередь это электро$ воздействия на семена и сами растения, осу$ ществляемые путем подведения напряжения к любой из частей системы почва–растение. Зна$ ние величин электротропизма поможет создавать наиболее комфортные по величине и направле$ нию электрические разности потенциалов для роста растений при электростимулировании, электроукоренении и ускоренном размножении растений, например, одревесневшими черенками [17, 19]. Кроме того, на основе знаний электро$ тропизма в системе почва–растение возможна оценка состояния дендрофлоры [13]. Это только первые сведения об обнаруженном явлении. По мере развития исследований в этом направлении представления об электропизме в системе почва– растение будут уточняться и находить все новые области практического применения.
ВЫВОДЫ
1. Независимо от вида растения и почвы, на которой оно произрастает, и каких бы то ни было других причин, электрический потенциал между почвой и растением всегда отрицательный: у поч$ вы больше, у растения меньше. Электрический потенциал травянистых растений в сильной мере зависит и от площади поверхности листа. У таких
ПОЧВОВЕДЕНИЕ № 7 2013