Материал: В-Л-Быков-Цитогогия_и_общая_гистология
рые клетки становятся двуядерными. Способность кардиомиоцитов человека к полному митотическому делению утрачивается к моменту рождения или в первые месяцы жизни. Вместе с тем. в этих клетках начинаются процессы полиплоидизации, протекающие, как предполагают, путем обычного, но незавершенного митоза и продолжающиеся в кардиомиоцитах желудочков до 8- 12 лет. Выстраиваясь в цепочки, сердечные миоциты не сливаются друг с другом (как это происходит при развитии скелетного мышечного волокна), а формируют сложные межклеточные соединения - вставочные диски, связывающие кардиомиоциты в функциональные волокна.
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ СЕРДЕЧНОЙ МЫШЕЧНОЙ ТКАНИ
Сердечная мышечная ткань образована клетками - кардиомиоцитами (сердечными миоцитами), связанными друг с другом в области вставочных дисков и образующими трехмерную сеть ветвящихся и анастомозирующих функциональных волокон (рис. 13-11).
Рис. 13-11. Сердечная мышечная ткань Клетки сердечной мышечной ткани - кардиомиоциты (КМЦ) - связаны друг с другом в области вставочных дисков (ВД) и благодаря наличию анастомозов (А) и разветвлений образуют трехмерную сеть функциональных волокон. Ядро (Я) занимает в КМЦ центральное положение, миофибриллы (МФ) располагаются по периферии, под сарколеммой (СЛ). КРС - кровеносные сосуды в прослойках соединительной ткани.
- 431 -
КАРДИОМИОЦИТЫ
Кардиомиоциты - цилиндрические или ветвящиеся клетки, более крупные в желудочках, где их длина составляет 100-150 мкм, а диаметр - 10 - 20 мкм. В предсердиях они обычно имеют неправильную форму и меньшие размеры (длина - 40-70 мкм, диаметр - 5-6 мкм). Кардиомиоциты содержат одно или два ядра и саркоплазму, покрыта сарколеммой, которая снаружи окружена базальной мембраной.
Ядра кардиомиоцитов - светлые, с преобладанием эухроматина, хорошо заметными ядрышками - занимают в клетке центральное положение. У взрослого человека (как и у всех исследованных до настоящего времени млекопитающих) более половины кардиомиоцитов являются двуядерными. Для кардиомиоцитов типична полиплоидия (более выраженная в желудочках), лишь часть из них являются диплоидными (виды с полностью диплоидными сердечными миоцитами не найдены). Степень полиплоидизации кардиомиоцитов характеризуется существенными индивидуальными различиями и даже у молодых здоровых мужчин варьирует в три раза. Предполагают, что степень полиплоидизации кардиомиоцитов у данного индивидуума является важным фактором, определяющим потенциальную способность его сердечной мышцы адаптироваться к повышенным нагрузкам.
Саркоплазма кардиомиоцитов содержит органеллы и включения, которые образуют следующие аппараты: 1) сократительный, 2) передачи возбуждения (с сарколеммы на сократительный аппарат), 3) опорный, 4) энергетический, 5) синтетический, 6) лизосомальный (аппарат внутриклеточного переваривания).
Сократительный аппарат сильно развит в сократительных (рабочих) кардиомиоцитах (в особенности, в желудочковых), которых он занимает до 5070% объема клетки. Слабое развитие этого аппарата свойственно проводящим и секреторным кардиомиоцитам (см. ниже). Сократительный аппарат кардиомиоцитов сходен с таковым в скелетных мышечных волокнах и также представлен миофибриллами, обладающими поперечной исчерченностью (средняя длина саркомера равна примерно 2 мкм). Вместе с тем, миофибриллы кардиомиоцитов нередко частично сливаются друг с другом (рис. 13-12), образуя единую структуру, а их сократимые белки биохимически отличаются от таковых в скелетной мышечной ткани. В саркоплазме кардиомиоцитов миофибриллы ориентированы продольно и располагаются по ее периферии, под сарколеммой.
- 432 -
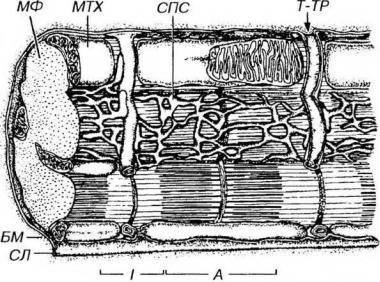
Рис. 13-12. Саркотубулярная система кардиомиоцита. Саркотубулярная система представлена саркоплазматической сетью (СПС), не образующей терминальных цистерн, и широкими Т-трубочками (Т-ТР). Обозначения компонентов саркомера - те же, что на рис. 13-3. Обратите внимание на частичное слияние МФ друг с другом в единую систему.
Аппарат передачи возбуждения (саркотубулярная система) в
кардиомиоцитах в целом сходен с таковым в скелетных мышечных волоконах, однако он обладает рядом особенностей.
Саркоплазматическая сеть развита слабее, чем в скелетном мышечном волокне, менее активно накапливает Са2+, не образует терминальных цистерн. Во время расслабления она выделяет ионы Са2+ в саркоплазму с низкой скоростью, что обеспечивает автоматизм кардиомиоцитов.
Поперечные (Т-) трубочки - широкие, содержат компоненты базальной мембраны, вместе с элементами саркоплазматической сети образуют диады (включают одну Т-трубочку и одну цистерну сети), которые располагаются в области Z-линий. Т-трубочки хорошо выражены в миоцитах желудочков и почти не обнаруживаются в предсердных миоцитах. Ионы Са2+ проникают в саркоплазму кардиомиоцитов не только из саркоплазматической сети, но также через Т-трубочки и сарколемму из межклеточного пространства.
- 433 -
Опорный аппарат кардиомиоцитов представлен элементами цитоскелета, обеспечивающими упорядоченное расположение миофиламентов и миофибрилл внутри волокна, а также базальной мембраной и сарколеммой. Его структурная и биохимическая организация сходна с таковой в волокнах скелетной мышечной ткани. В кардиомиоцитах его особенностью служит то, что элементы цитоскелета связаны с особыми межклеточными соединениями - вставочными дисками (см. ниже). Сарколемма содержит интегрины - трансмембранные гликопротеины, которые опосредуют связь элементов цитоскелета кардиомиоцитов с компонентами межклеточного вещества (коллагеном, ламинином и фибронектином). Снаружи сарколемма кардиомиоцитов окружена базальной мембраной, в которую вплетаются ретикулярные и тонкие коллагеновые волокна.
Вставочные диски осуществляют связь кардиомиоцитов друг с другом.
Под световым микроскопом они имеют вид поперечных прямых или зигзагообразных полосок, пересекающих функциональное волокно сердечной мышечной ткани (см. рис. 13-11). Под электронным микроскопом определяется сложная организация вставочного диска, представляющего собой комплекс межклеточных соединений нескольких типов (рис. 13-13). В области поперечных (ориентированных перпендикулярно расположению миофибрилл) участков вставочного диска соседние кардиомиоциты образуют многочисленные интердигитации, связанные контактами типа десмосом и полосок слипания (fasciae adherentes). Актиновые филаменты прикрепляются к поперечным участкам сарколеммы вставочного диска на уровне Z-полоски. В области вставочного диска выявляются гликопротеины кадгерины, которые обеспечивают Са-зависимое адгезивное соединение кардиомиоцитов друг с другом. На сарколемме продольных участков вставочного диска имеются многочисленные щелевые соединения, обеспечивающие ионную связь кардиомиоцитов и передачу импульса сокращения.
Энергетический аппарат кардиомиоцитов представлен митохондриями и включениями, расщепление которых обеспечивает получение энергии. Митохондрии лежат рядами между миофибриллами, у полюсов ядра и под сарколеммой. Они очень многочисленные и крупные (занимают около 35-40% объема саркоплазмы - значительно больше, чем в волокнах скелетной мышечной ткани), с плотно расположенными поперечными кристами (см. рис. 13-12), что в совокупности отражает высокий уровень дыхательной активности сердечной мышечной ткани. Митохондрии кардиомиоцитов обладают также свойством накапливать кальций в высоких концентрациях.
- 434 -
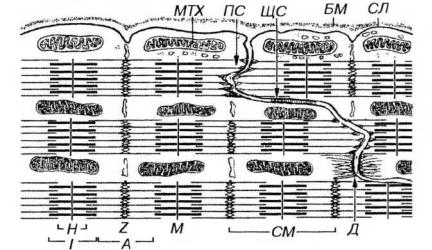
Рис. 13-13. Ультраструктурная организация области вставочного диска кардиомиоцитов. В поперечных участках вставочного диска соседние кардиомиоциты образуют многочисленные интердигитации, связанные контактами типа десмосом (Д). Актиновые филаменты прикрепляются к поперечным участкам сарколеммы вставочного диска в участке полоски слипания (ПС) На сарколемме продольных участков вставочного диска располагаются щелевые соединения (ЩС). БМ - базальная мембрана, СЛ - сарколемма, МТХ - митохондрия. Обозначения компонентов саркомера (СМ) - те же, что на рис. 13-3.
Энергия, необходимая кардиомионитам, получается ими путем расщепления основного энергетического субстрата этих клеток - жирных кислот, переносимых в саркоплазму в форме липопротеинов. Жирные кислоты депонируются в виде триглицеридов в многочисленных липидных каплях, содержание которых значительно выше, чем в скелетных мышцах. Липидные капли располагаются преимущественно у полюсов ядра. Другой энергетический субстрат - гликоген - находится в виде гранул, расположенных преимущественно между миофибриллами и миофиламентами, главным образом, в области I-диска. Кардиомиоциты. как и волокна скелетной мышечной ткани, характеризуются наличием в их саркоплазме железосодержащего кислород-связывающего пигмента миоглобина, придающего им красный цвет и сходного по строению и функции с гемоглобином эритроцитов.
Синтетический аппарат кардиомиоцитов в сократительных (рабочих)
кардиомиоцитах выражен умеренно; он значительно развит лишь в секреторных кардиомиоцитах (см. ниже). Синтетический аппарат включает свободные рибосомы и полисомы, цистерны грЭПС
- 435 -