Материал: В-Л-Быков-Цитогогия_и_общая_гистология
ности, отмечается в биоптатах мышц при различных миопатнях, при которых процессы повреждения сочетаются с явлениями регенерации.
Полноценная регенерация мышечных волокон возможна лишь при их незначительных дефектах; ее условием является сохранение целостности их базальной мембраны. Предполагают, что базальная мембрана служит своеобразным барьером, предотвращающим проникновение клеток фибробластического ряда в поврежденное волокно, но пропускающим макрофаги, поглощающие некротизированную ткань. Она осуществляет также роль направляющей, поддерживающей и ориентирующей структуры для мигрирующих миосателлитоцитов и для формирующихся мышечных трубочек, обеспечивает условия микроокружения, оптимальные для процесса регенерации.
Неполноценная регенерация мышечных волокон наблюдается при значительной травме мышцы (сопровождающейся обширным повреждением не только мышечных волокон, но и соединительнотканных структур). Полноценной регенерации в этих случаях обычно препятствует разрастание соединительной ткани эндо- и перимизия (см. ниже). Последняя быстро заполняет область дефекта и в конечном итоге образует в области краев поврежденных мышечных волокон соединительнотканный рубец, который является барьером, препятствующим их воссоединению. Функция мышцы при этом остается нарушенной.
Эктопическое развитие костной ткани внутри поврежденной скелетной мышцы отмечается в некоторых случаях через 2-3 нед. после травмы. Оно происходит, очевидно, вследствие активизации малодифференцированных остеогенных клеток-предшественников, лежащих в соединительнотканных структурах травмированного участка мышцы, которые превращаются в остеобласты. Индукции этих клеток метут способствовать вещества, выделяемые поврежденными мышечными волокнами.
СКЕЛЕТНАЯ МЫШЦА КАК ОРГАН
Скелетная мышца состоит из пучков мышечных волокон, связанных воедино системой соединительнотканных компонентов.
Количество мышечных волокон в отдельных мышцах человека варьирует в широких пределах. Так, в четырехглавой мышце бедра (m. quadriceps femoris) оно составляет около 1.7 млн., икроножной мышце (m. gastrochnemius) - 1-1.5 млн., портняжной мышце (m. sarto-
- 426 -
rius) - 100-200 тыс., в двуглавой и трехглавой мышцах плеча (m. biceps и т. triceps brachii) оно примерно одинаково и колеблется в пределах 200-600 тыс.
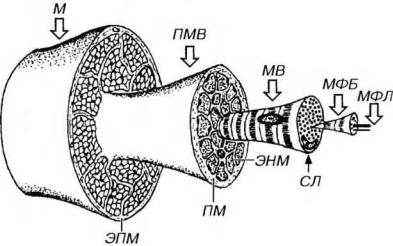
Соединительнотканные компоненты мышцы представлены эпимизием,
перимизием и эндомизием (рис. 13-10). Общее содержание соединительной ткани в скелетной мышце очень значительно - в зависимости от типа мышцы коллаген соединительной ткани составляет от 3 до 30% ее белков.
Рис. 13-10. Структурная организация скелетной мышцы. Мышца (М) покрыта эпимизием (ЭПМ), отдающим вглубь более тонкие соединительнотканные перегородки - перимизий (ПМ), который образует оболочки пучков мышечных волокон (ПМВ). От ПМ внутрь ПМВ отходят тончайшие прослойки рыхлой волокнистой соединительной ткани, окружающие каждое мышечное волокно (МВ), именуемые эндомизием (ЭНМ). МВ покрыто сарколеммой (СЛ) и базальной мембраной (не показана) и обладает поперечной исчерченностью; в его центральной части содержатся поперечно исчерченные миофибриллы (МФБ), образованные системой тонких и толстых миофиламентов - МФЛ.
Эпимизий - тонкий, прочный и гладкий снаружи чехол из плотной волокнистой соединительной ткани, окружающий всю мышцу.
Перимизий - тонкие, разветвляющиеся и не всегда четко очерченные соединительнотканные перегородки, отходящие от внутренней поверхности эпимизия вглубь мышцы. Он образует оболочки отдельных пучков мышечных волокон, численностью 10-100 (наиболее часто - около 20) волокон.
- 427 -
Эндомизий - тончайшие прослойки рыхлой волокнистой соединительной ткани, отходящие от перимизия внутрь пучков мышечных волокон и окружающие каждое мышечное волокно. Соединительнотканные волокна эндомизия вплетаются в базальную мембрану мышечных волокон.
КРОВОСНАБЖЕНИЕ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ
Кровоснабжение скелетной мышечной ткани осуществляется артериями, которые проникают в мышцу вместе с нервами через эпимизий и идут вдоль прослоек перимизия, постепенно разветвляясь. Между этими ветвями имеются многочисленные анастомозы. Тонкие артериальные веточки располагаются перпендикулярно длинной оси мышечных волокон, а отходящие от них капилляры проходят в эндомизии вдоль волокон, соединяясь перемычками и образуя густую сеть. При сокращении мышцы капилляры спиралеобразно скручиваются. Мышца относится к числу обильно васкуляризованных тканей - на одно мышечное волокно приходится в среднем 3-4 капилляра.
ИННЕРВАЦИЯ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ
Скелетные мышцы обладают эфферентной (двигательной) и афферентной (чувствительной) иннервацией.
Эфферентная иннервация скелетных мышц обеспечивается нервными волокнами (аксонами α-мотонейронов), образующими на мышечных волокнах специализированные нервно-мышечные окончания (нервно-мышечные синапсы, или моторные бляшки), которые осуществляют передачу возбуждения с нервного волокна на мышечное. Описание нервно-мышечных окончаний представлено в главе 14. Один мотонейрон может иннервировать различное количество мышечных волокон.
Двигательная единица (ДЕ) представляет собой совокупность мотонейрона и иннервируемых им мышечных волокон. Количество мышечных волокон, входящих в одну ДЕ, минимально в мелких мышцах, осуществляющих точные и тонкие движения. В глазных мышцах человека ДЕ включает от 2-6 до 13-20 мышечных волокон, в мышцах кисти - от 10-25 до 100-300, в мышцах туловища - 1500-2000. Управление мышечной активностью обеспечивается как изменением частоты активации ДЕ, так и вовлечением различного их числа в процесс сокращения.
- 428 -
Мышечные волокна, образующие одну ДЕ, обладают одинаковыми механическими свойствами, гистохимическими характеристиками и относятся к одному типу, однако рассеяны по обширной территории мышцы.
Афферентная иннервация скелетных мышц обеспечивается нервномышечными веретенами - рецепторами растяжения волокон поперечнополосатых мышц, которые представляют собой сложные инкапсулированные нервные окончания, состоящие из веточек нервных волокон, оплетающих особые тонкие (интрафузальные) мышечные волокна, заключенные в тончайшую соединительнотканную капсулу. Остальные мышечные волокна называются экстрафузальными. Нервно-мышечные веретена описаны в главе 14.
Клиническое значение нарушений структурно-функциональной организации скелетных мышц. Система скелетных мышц поражается разнообразными заболеваниями, из которых наибольшее клиническое значение имеют две группы. Первая включает тяжелые дистрофические расстройства (часто генетически обусловленные), при которых первично нарушается структура и функция мышечной ткани. Вторая группа заболеваний обусловлена нарушением иннервации мышечных волокон.
Мышечная дистрофия Дюшенна является наиболее распространенным заболеванием, относящимся к первой группе и имеющим наследственный характер. Она поражает мальчиков, проявляется нарастающей мышечной слабостью и приводит к смерти в молодом возрасте. Причина заболевания заключается, по-видимому, в нарушении функции гена, контролирующего выработку белка дистрофина, функция которого, как предполагают, заключается в обеспечении связи между миофибриллами и элементами межклеточного вещества (см. выше). В отсутствие этого белка мышечные волокна становятся очень непрочными, легко повреждаются и гибнут при небольших нагрузках, замещаясь соединительной тканью.
Амиотрофический латеральный склероз представляет собой наследственное заболевание, относящееся ко второй из указанных групп. При этом заболевании атрофия мышечной ткани и смерть больного от поражения дыхательных мышц обусловлены дегенеративными изменениями мотонейронов спинного мозга.
Злокачественная миастения (myasthenia gravis) также относится к мышечным расстройствам, обусловленным нарушениями иннервации. В ее основе лежит аутоиммунный процесс, который характеризуется образованием антител к рецепторам ацетилхолина на сарколемме нервно-мышечного синапса. Связывание этих антител с рецепторами приво-
- 429 -
дит к нарушению их функции. При миастении отмечается прогрессирующая резко выраженная мышечная слабость.
Ботулизм является пищевым отравлением токсином, вырабатываемым бактериями Clostridium botulinum, который нарушает выделение медиатора в нервно-мышечном синапсе. Это тяжелое заболевание сопровождается параличом скелетных мышц.
СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ
Сердечная мышечная ткань (поперечнополосатая мышечная ткань целомического типа) встречается только в мышечной оболочке сердца (миокарде) и устьях связанных с ним крупных сосудов. Ее клетки (сердечные миоциты, или кардиомиоциты) составляют лишь 30-40% общего числа клеток сердца, но образуют 70-90% его массы. Основным функциональным свойством сердечной мышечной ткани служит способность к спонтанным ритмическим сокращениям, на активность которых влияют гормоны и нервная система (симпатическая и парасимпатическая).
ГИСТОГЕНЕЗ СЕРДЕЧНОЙ МЫШЕЧНОЙ ТКАНИ
Источником развития сердечной мышечной ткани служит миоэпикардиальная пластинка висцерального листка спланхнотома (целомическая выстилка в шейной часта эмбриона). Клетки этой пластинки (миобласты) активно размножаются митозом и постепенно образуют миофиламенты, формирующие миофибриллы. С появлением последних клетки именуются сердечными миоцитами, или кардиомиоцитами). Миофибриллы первоначально не обладают поперечной исчерченностыо и строгой ориентацией в клетке; в дальнейшем они располагаются вдоль ее длинной оси, а их тонкие филаменты прикрепляются к уплотненным участкам сарколеммы (Z-вешеству) у концов кардиомиоцитов.
Дифференцировка кардиомиоцитов, в отличие от волокон скелетной мышечной ткани, сочетается с их размножением: гликоген и миофибриллы накапливаются в саркоплазме клеток, которые еще продолжают делиться, уже обладая сократительной способностью. В период деления сердечных миоцитов часть их миофибрилл подвергается распаду с последующей повторной сборкой. В цитоплазме дифференцирующихся кардиомиоцитов нарастает содержание рибосом, цистерн грЭПС, митохондрий. Из-за отсутствия цитотомии при делении некото-
- 430 -