Материал: В-Л-Быков-Цитогогия_и_общая_гистология
Расслабление после мышечного сокращения происходит в результате снижения концентрации Са2+ в области саркомера, которое вызывает отщепление Са2+ от ТnС-субъединицы тропонина и возвращение тропонина в первоначальное конформационное состояние. Нити тропомиозина при этом вновь закрывают активные центры на молекулах актина, что обусловливает прекращение циклического образования мостиков.
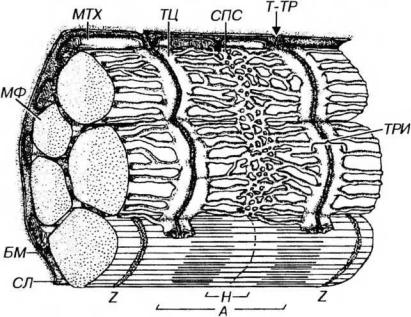
Аппарат передачи возбуждения (саркотубулярная система) необходим для того, чтобы распространяющаяся по сарколемме волна деполяризации могла вызвать срабатывание сократительного аппарата миофибрилл. В мышечном волокне связь между возбуждением и сокращением выполняют две специализированные мембранные системы - саркоплазматическая сеть и поперечные (Т-) трубочки (от англ. transverse - поперечный), образующие функционально единую саркоту-булярную систему (рис. 13-8).
Рис. 13-8. Саркотубулярная система волокна скелетной (соматической) мышечной ткани. Саркотубулярная система включает саркоплазматическую сеть (СПС) и поперечные, или Т-трубочки (Т-ТР). СПС окружает каждый саркомер миофибриллы; ее трубочки сливаются, образуя пары плоских терминальных цистерн (ТЦ). Т-ТР представляют собой впячивания сарколеммы (С/1), отходящие от нее под прямым углом и проникающие в промежуток между двумя ТЦ, в совокупности с которыми они формируют триады (ТРИ). МФ - миофибриллы. Обозначения компонентов саркомера - те же, что на рис.
13-3.
- 416 -
Саркоплазматическая сеть - система уплощенных, вытянутых и анастомозирующих мембранных трубочек и мешочков, которая окружает каждый саркомер миофибриллы наподобие муфты. В области наружных отделов А- и I-дисков трубочки сливаются, образуя пары плоских терминальных цистерн (на каждый саркомер приходится по две такие пары). Саркоплазматическая сеть обладает выраженной способностью депонировать и выделять ионы кальция. Ее мембрана содержит высокие концентрации интегральных белков, являющихся кальциевыми насосами, а на внутренней поверхности находится белок кальсеквестрин, связывающий ионы Са2+.
Поперечные (Т-) трубочки представляют собой впячивания сарколеммы, отходящие от нее под прямым углом к оси волокна и расположенные у млекопитающих вблизи границы I- и А- дисков. Ветви соседних Т-трубочек опоясывают каждый саркомер и анастомозируют друг с другом. Конечные участки Т-трубочек проникают в промежуток между двумя терминальными цистернами саркоплазматической сети (см. рис. 13-8), формируя вместе с ними особые структуры - триады. В области триады между параллельно лежащими мембранами Т-трубочки и терминальных цистерн, разделенными узкой щелью, имеются специализированные контакты, которые образованы рядами плотных частиц (ножек), предположительно служащие каналами выделения кальция.
Выделение кальция происходит после того, как волна деполяризации с поверхности сарколеммы по Т-трубочкам распространяется вглубь волокна. В области триад возбуждение передается на мембрану саркоплазматической сети и вызывает повышение ее проницаемости. Это приводит к быстрому выделению из ее элементов ионов кальция (главным образом, в области терминальных цистерн). Выделившийся Са2+ диффундирует в миофибриллы, где он, присоединяясь к тропониу, запускает механизм взаимодействия актина и миозина (см. выше). Концентрация Са2+ вокруг миофиламентов при этом резко повышается с 10-7М до 10-5М.
Активный обратный транспорт кальция в саркоплазматическую сеть
(секвестрация кальция) происходит наряду с его выбросом, который представляет собой кратковременный процесс. Обратный транспорт Са2+ осуществляется благодаря деятельности кальциевых насосов (Са-зависимой АТФазы) в мембране саркоплазматической сети. Падение концентрации Са2+ вследствие его секвестрации приводит к возвращению тропонина в первоначальное конформационное состояние, прекращению взаимодействия миозиновых мостиков с актином и расслаблению мышечного волокна.
- 417 -
Опорный аппарат мышечного волокна включает особые элементы цитоскелета, обеспечивающие высокоупорядоченное расположение миофиламентов и миофибрилл внутри волокна, а также связанную с ними сарколемму и базальную мембрану (см. рис. 13-3, 13-6 и 13-8), соединяющие мышечное волокно с сухожилием, на которое передается усилие, развиваемое волокном при сокращении.
Телофрагма (Z-линия) - область прикрепления тонких миофиламентов двух соседних саркомеров; она имеет вид плотной полоски шириной 30-100 нм без резких границ. Представляет собой сложную трехмерную решетку из особых тонких нитей (Z-филаментов), идущих зигзагообразно под углом 45° к оси саркомера и образующих тетрагональную (четырехугольную) структуру, связывающую тонкие нити двух соседних саркомеров. В ячейках решетки этих филаментов имеется плотный материал. В состав Z-линий входит ряд белков: α- актиния, филамин, Z-белок.
Мезофрагма (М-линия) - плотная линия шириной 75-85 нм, расположенная в центре А-диска и являющаяся областью закрепления толстых (миозиновых) филаментов в саркомере. Она образована центральными участками миозиновых филаментов, которые располагаются в виде гексагональных фигур и связаны друг с другом системой мостиков, состоящих из тонких нитей белков миомезина, креатинкиназы и М-белка.
Титин (коннектин) представляет собой белок с эластическими свойствами, нити которого присоединены к толстым филаментам по всей их длине и, продолжаясь в I-диски, прикрепляют концы толстых филаментов к Z- линиям. Таким образом, нити титина связывают М и Z-линии, и, благодаря своей эластичности, препятствуют перерастяжению мышцы. Они образуют внутри саркомера решетчатую структуру и поддерживают упорядоченное взаимное расположение системы толстых и тонких миофиламентов.
Небулин - белок, имеющий вид удлиненных нитей, расположенных по всей ширине I-диска параллельно тонким филаментам, с которыми он связан. Предполагается, что небулин отвечает за поддержание длины тонких филаментов и (или) обеспечивает их механическую стабилизацию.
Промежуточные филаменты (диаметром около 10 нм), состоящие из белка десмина, являются важным элементом цитоскелета и образуют в пределах мышечного волокна две пространственные системы. Первая
- 418 -
состоит из филаментов, которые располагаются в саркомерах продольно и связывают соседние телофрагмы одной миофибриллы. Вторая представлена поперечно ориентированными филаментами, которые связывают мезофрагмы, а также телофрагмы соседних миофибрилл друг с другом. Такие же филаменты прикрепляют телофрагмы к сарколемме и элементам системы Т-трубочек и сарконлазматической сети. Благодаря описанной организации системы промежуточных филаментов поддерживается упорядоченное взаимное расположение саркомеров соседних миофибрилл и других компонентов мышечного волокна.
Дистрофии - белок, одними участками прикрепляющийся к актиновым филаментам, а другими - к комплексу гликопротеинов, которые пронизывают сарколемму и связываются на ее поверхности с компонентами базальной мембраны. Таким путем усилия, создаваемые внутри мышечного волокна, посредством ряда белков передаются на элементы межклеточного вещества. Генетический дефект, связанный с нарушением выработки дистрофина, обусловливает развитие мышечного заболевания - дистрофии Дюшенна (см. ниже).
Костамеры - кольца из белка винкулина, охватывающие изнутри мышечное волокно и расположенные перпендикулярно к его длинной оси. Они представляют собой участки непосредственного соединения между сарколеммой и подлежащими I-дисками миофибрилл. Благодаря наличию в костамерах интегринов они, также, возможно, являются структурами, которые через адгезивный гликопротеин фибронектин связывают элементы межклеточного вещества (например, коллагеновые волокна) с миофибриллами. Помимо винкулина, в костамерах имеются другие белки, связанные с цитоскелетом: талин, сиектрин, α-актинин.
Структура краевых участков мышечных волокон. На концах мышечных волокон сарколемма, покрытая базальной мембраной, образует многочисленные глубокие впячивания, в которые вдаются коллагеновые волокна сухожилия, вплетающиеся в базальную мембрану и прочно связывающие сухожилие с мышечными волокнами.
Энергетический аппарат мышечных волокон представлен митохондриями, вырабатывающими энергию, необходимую для осуществления мышечной работы, синтетических, транспортных и других процессов жизнеобеспечения, а также трофическими включениями, содержащими вещества, расщепление которых служит источником энергии.
Митохондрии в миосимпласте располагаются в виде цепочек под сарколеммой и между миофибриллами (см. рис. 13-8). Они имеют вытя-
- 419 -
нутую форму, содержат большое количество поперечно расположенных ламеллярных крист, характеризуются высокой активностью окислительновосстановительных ферментов. Их содержание и размеры больше в красных волокнах, чем в белых (см. ниже) и увеличиваются при тренировке мышц.
Энергия, необходимая для осуществления мышечной работы, запасается в мышечных волокнах в виде АТФ и фосфокреатина - энергоемких фосфатных соединений. Источником энергии служит расщепление гликогена и липидов. При кратковременных резких нагрузках на скелетные мышцы источником энергии служит глюкоза, получаемая преимущественно в результате расщепления гликогена. Главным источником энергии при выполнении работы, требующей выносливости, служат жирные кислоты.
Гликоген находится в саркоплазме (преимущественно белых волокон - см. ниже) в виде β-частиц диаметром 20-30 нм. Последние образуют скопления между миофибриллами, большей частью на уровне I-дисков. Запасы гликогена, составляющие 0.5-1% массы волокна, опустошаются при длительной интенсивной нагрузке.
Липидные капли располагаются между миофибриллами по всей толщине миосимпласта, образуя скопления преимущественно на уровне I-дисков. Их содержание варьирует в широких пределах, но в среднем выше в красных волокнах (0.5% объема саркоплазмы), чем в белых (0.2%).
Миоглобин - железосодержащий кислород-связывающий пигмент мышечных волокон, придающий им красный цвет и сходный по строению и функции с гемоглобином эритроцитов - типичное включение мышечного волокна, которое можно условно отнести к энергетическому аппарату. Миоглобин находится в более высоких концентрациях в красных волокнах (что и определяет их цвет); его способность к связыванию кислорода способствует повышению активности процессов окислительного фосфорилирования).
Синтетический аппарат мышечного волокна представлен свободными рибосомами и полирибосомами (особенно многочисленными под сарколеммой в области I-диска и вблизи ядер), цистернами грЭПС и комплексом Гольджн, элементы которого в виде сотен или тысяч стопок мешочков рассеяны по саркоплазме миосимпласта.
Лизосомальный аппарат (аппарат внутриклеточного переваривания)
в мышечных волокнах необходим для обеспечения постоянно протекающего процесса обновления его структурных
- 420 -