Материал: В-Л-Быков-Цитогогия_и_общая_гистология
ненные зеркально концами друг с другом в области М-линии, формируют толстые нити с центральной гладкой частью длиной около 0.2 мкм и двумя периферическими участками, в которых от центрального стержня отходят миозиновые головки (около 500). Миозин головок обладает АТФазной активностью (способностью осуществлять гидролиз АТФ), однако в отсутствие его взаимодействия с актином скорость гидролиза АТФ ничтожно мала.
Рис. 13-4. Строение толстых миофиламентов (по К.Де Дюву, 1987, с изменениями). 1 - молекула миозина имеет вид нити с двумя головками (Г) на одном конце. Миозин включает легкий меромиозин (ЛММ), образующий стержневую часть молекулы, и тяжелый меромиозин (ТММ), соответствующий участкам Г и связующей шейки. Участки сгибания молекулы миозина показаны стрелками. 2 - стержневые части молекул миозина собраны в пучки (П), снаружи которых располагаются миозиновые Г. 3 - толстые миофиламенты (ТЛМФ) образованы П молекул миозина, соединенными зеркально концами друг с другом. Центральная часть ТЛМФ - гладкая, периферические содержат многочисленные миозиновые Г.
Тонкие нити (миофиламенты) содержат сократимый белок актин (на него приходится 20% белков миофибриллы) и два регуляторных белка - тропонин (около 2%) и тропомиозин (около 7%). Последние формируют функционально единый тропонин-тропомиозиновый комплекс.
Актин в мономерной форме представлен полярными глобулярными субъединицами диаметром 4-5 нм (G-актин), которые имеют активные центры, способные связываться с молекулами миозина. G-актин агрегирует с образованием полимерного фибриллярного актина (F-актина), молекула которого имеет вид двух скрученных нитей толщиной 7 нм и вариабельной длины (рис. 13-5).
- 411 -
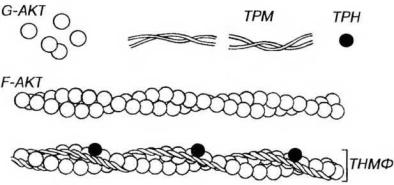
Рис. 13-5. Строение тонких миофиламентов. Тонкие миофиламенты (ТНМФ) содержат сократимый белок актин (АКТ) и два регуляторных белка - тропонин (TPH) и тропомиозин (TPM). Глобулярные субъединицы АКТ (G-AKT), агрегируют с образованием фибриллярного АКТ (F-AKT), молекула которого имеет вид двух скрученных нитей. TPM образован нитевидными молекулами, соединяющимися своими концами и образующими тяж, лежащий в борозде молекулы F-AKT. ТРН - глобулярный белок, связанный с молекулой ТРМ и формирующий с ней функционально единый комплекс ТРН-ТМ.
Тропомиозин представлен нитевидными молекулами, которые соединяются своими концами, образуя длинный тонкий тяж, лежащий в борозде, образуемой перевитыми нитями F-актина. Так как таких борозд на молекуле актина две, то и тропомиозиновых нити тоже две. Всего в состав тонкой нити входит примерно 50 молекул тропомиозина.
Тропонин представляет собой глобулярный белок, каждая его молекула располагается на тропомиозиновой молекуле вблизи ее конца. Тропонин состоит из трех субъединиц: ТnС - связывающей кальций, ТnТ - прикрепляющейся к тропомиозину, и TnI - ингибирующей связывание миозина с актином.
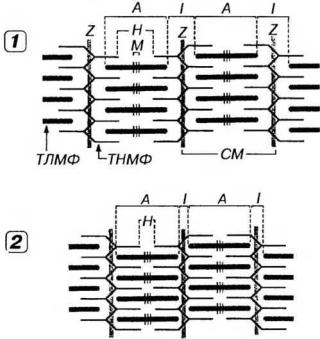
Механизм мышечного сокращения описывается теорией скользящих нитей, согласно которой укорочение каждого саркомера (а, следовательно, миофибрилл и всего мышечного волокна) при сокращении происходит благодаря тому, что тонкие нити вдвигаются в промежутки между толстыми без изменения их длины (рис. 13-6).
Скольжение нитей в саркомере и усилие, развиваемое мышцей, обеспечиваются благодаря циклической активности миозиновых мостиков, которые при сокращении повторно прикрепляются к актину, обес-
- 412 -
печивают усилие тяги, а затем открепляются от него (рис. 13-7). В этом механизме АТФ играет двойную роль, обеспечивая энергию, необходимую как для осуществления сокращения, так и для открепления мостиков.
Рис. 13-6. Механизм мышечного сокращения в соответствии с теорией скользящих нитей. Укорочение саркомеров (СМ) при сокращении (2) по сравнению с их состоянием в покое (1) происходит благодаря тому, что тонкие миофиламенты (ТНМФ) вдвигаются в промежутки между толстыми (ТЛМФ) без изменения их длины. Остальные обозначения - как на рис. 13-3.
Строгая пространственная упорядоченность взаимодействия множества толстых и тонких нитей в саркомере определяется наличием сложно организованного поддерживающего аппарата (см. ниже). Его элемента на всех этапах мышечного сокращения и расслабления, динамично перестраиваясь, фиксируют и удерживают миофиламенты в правильном положении, которое оптимальным образом обеспечивает их взаимный контакт, взаимодействие и взаимное скольжение.
- 413 -
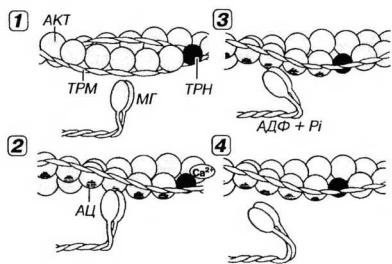
Рис. 13-7. Молекулярные механизмы мышечного сокращения. 1 - в покое миозиновые головки (МГ), с которыми связаны молекулы АТФ, неспособны взаимодействовать с активными центрами (АЦ) на молекуле актина (АКТ), потому что последние прикрыты комплексом тропонин-тропомиозин (ТРНТРМ). 2 - мышечное сокращение начинается вследствие повышения концентрации Са2+, который воздействует на ТРН. Возникающее изменение конформации ТРН и смещение молекулы связанного с ним ТРМ демаскирует АЦ на молекуле АКТ, с которыми связываются МГ, образуя поперечные мостики. 3 - за счет сгибания МГ в области их прикрепления к молекуле АКТ развивается усилие, смещающее тонкие миофиламенты (ТНМФ) вдоль толстых (ТЛМФ) к центру саркомера (см. рис. 13-06). АТФ при этом гидролизуется до АДФ и фосфата (Pi). 4 - размыкание мостика и его отделение от ТНМФ наступают вследствие связывания с ним новой молекулы АТФ. Далее мостик принимает исходное положение (перпендикулярное ТНМФ) и начинается новый цикл сокращения. Циклическое взаимодействие МГ и ТНМФ будет продолжаться при сохранении высокой концентрации ионов Са2+ и наличии АТФ.
В покое (при очень низкой концентрации ионов Са2+) в миофибрилле расслабленного мышечного волокна толстые и тонкие нити не соприкасаются. Миозиновые головки (с которыми связаны молекулы АТФ) не могут взаимодействовать с активными центрами (участками связывания миозина) на молекуле актина, потому что последние прикрыты тропонин-тропомиозиновым комплексом. Толстые и тонкие филаменты беспрепятственно скользят друг относительно друга. При этом мышечные волокна почти не сопротивляются пассивному растяжению. Такое состояние свойственно разгибательной мышце при сокращении соответствующей сгибательной. В отсутствие тропомиозина и тропонина (в условиях in vitro) миозин непрерывно взаимодействует с актином (пока имеется АТФ).
- 414 -
Мышечное сокращение вызывается резким повышением концентрации ионов Са2+ в области миофиламентов и включает несколько этапов (см. рис. 13-
7[2-4]).
A. Связывание ионов Са2+ с тропонином и освобождение активных центров на молекуле актина. Ионы Са2+ связываются с ТnС-субъединицами тропонина на тонких филаментах. При этом тропонин изменяет свою конформацию, смещает молекулы тропомиозина и открывает активные центры (участки связывания миозина) на молекуле актина.
Б. Связывание миозина и актина (формирование поперечных мостиков).
Миозиновые головки связываются с активными центрами на молекуле актина, формируя мостики, расположенные перпендикулярно продольной оси нити. Менее чем через 1 мс после этого под влиянием актомиозинового комплекса происходит гидролиз АТФ и отщепление его продуктов (АДФ и неорганического фосфата). При этом угол наклона мостика относительно продольной оси нити изменяется до 40°. Такой конформационный переход, происходящий в области прикрепления головки миозиновой молекулы, обусловливает развитие усилия и смещение тонких филаментов к центру саркомера. Предполагается, что "рабочий ход" миозинового мостика составляет около 10 нм; таким образом за один цикл мостик вызывает относительное перемещение тонких нитей на расстояние, равное примерно 1/200 длины саркомера.
B. Размыкание мостика. Связывание новой молекулы АТФ с мостиком вызывает его отделение от тонкого филамента. Мостик размыкается, возвращаясь в прежнее положение относительно миозиновой нити и может прийти в замыкание со следующим активным центром на тонкой. Каждый цикл замыкания-размыкания сопровождается расщеплением молекулы АТФ. В живой мышце это осуществляется с интервалом в несколько десятков миллисекунд после присоединения новой молекулы АТФ. В трупной мышце, где АТФ отсутствует, мостик не может разомкнуться, и мышца переходит в состояние трупного окоченения (rigor mortis).
При сокращении мышцы не происходит одновременного замыкания всех мостиков - их число нарастает по ходу его развития. При последующем расслаблении мышцы число мостиков снижается.
Изменение длины саркомера при сокращении является результатом относительного продольного смещения толстых и тонких нитей. При этом ширина A-диска не меняется; по мере проникновения в него тонких нитей происходит укорочение I-диска; соответственно значительно сужается Н- полоска (см. рис. 13-6).
- 415 -