Материал: t1_struktura_immunnoy_sistemy
Основные этапы активации системы комплемента.
Классический и альтернативный пути активации системы комплемента.
Система комплемента работает как биохимический каскад реакций. Комплемент активируется тремя биохимическими путями: классическим, альтернативным и лектиновым путем. Все три пути активации производят разные варианты C3-конвертазы (белка, расщепляющего С3). Классический путь (он был открыт первым, но эволюционно является новым) требует антител для активации (специфический иммунный ответ, приобретённый иммунитет), в то время как альтернативный и лектиновый пути могут быть активизированы антигенами без присутствия антител (неспецифический иммунный ответ, врождённый иммунитет). Итог активации комплемента во всех трёх случаях одинаков: C3-конвертаза гидролизует СЗ, создавая C3a и C3b и вызывая каскад дальнейшего гидролиза элементов системы комплемента и событий активации. В классическом пути для активации С3-конвертазы необходимо образование комплекса С4bC2a. Этот комплекс образуется при расщеплении С2 и С4 С1-комплексом. С1-комплекс, в свою очередь, для активации должен связаться с иммуноглобулинами класса М или G. C3b связывается с поверхностью болезнетворных микроорганизмов, что приводит к большей «заинтересованности» фагоцитов к связанным с СЗb клеткам (опсонизация). C5a — важный хемоаттрактант, помогающий привлекать в район активации системы комплемента новые иммунные клетки. И C3a, и C5a имеют анафилотоксическую активность, непосредственно вызывая дегрануляцию тучных клеток (как следствие — выделение медиаторов воспаления). C5b начинает формирование мембраноатакующих комплексов (МАК), состоящим из C5b, C6, C7, C8 и полимерного C9. МАК — цитолитический конечный продукт активации системы комплемента. МАК формирует трансмембранный канал, вызывающий осмотический лизис клетки-мишени. Макрофагипоглощают помеченные системой комплемента болезнетворные микроорганизмы.
Биологические функции
Сейчас выделяют следующие функции:
-
Опсонизирующая функция. Сразу вслед за активацией системы комплемента образуются опсонизирующие компоненты, которые покрывают патогенные организмы или иммунные комплексы, привлекая фагоцитов. Наличие на поверхности фагоцитирующих клеток рецептора к С3b усиливает их прикрепление к опсонизированным бактериям и активирует процесс поглощения. Такое более тесное прикрепление С3b-связанных клеток или иммунных комплексов к фагоцитирующим клеткам получило название феномена иммунного прикрепления.
-
Солюбилизация (т.е. растворение) иммунных комплексов (молекулой C3b). При недостаточности комплемента развивается иммунокомплексная патология (СКВ-подобные состояния). [СКВ = системная красная волчанка]
-
Участие в воспалительных реакциях. Активация системы комплемента приводит к выделению из тканевых базофилов (тучных клеток) и базофильных гранулоцитов крови биологически активных веществ (гистамина, серотонина, брадикинина), которые стимулируют воспалительную реакцию (медиаторов воспаления). Биологически активные компоненты, которые образуются при расщепленииС3 и С5, приводят к высвобождению вазоактивных аминов, таких как гистамин, из тканевых базофилов (тучных клеток) и базофильных гранулоцитов крови. В свою очередь это сопровождается расслаблением гладкой мускулатуры и сокращением клеток эндотелия капилляров, усилением сосудистой проницаемости. Фрагмент С5а и другие продукты активации комплемента содействуют хемотаксису, агрегации и дегрануляции нейтрофилов и образованию свободных радикалов кислорода. Введение С5а животным приводило к артериальной гипотонии, сужению легочных сосудов и повышению проницаемости сосудов из-за повреждения эндотелия. Фукнции С3а:
-
выступать в роли хемотаксического фактора, вызывая миграцию нейтрофилов по направлению к месту его высвобождения;
-
индуцировать прикрепление нейтрофилов к эндотелию сосудов и друг к другу;
-
активировать нейтрофилы, вызывая в них развитие респираторного взрыва и дегрануляцию;
-
стимулировать продукцию нейтрофилами лейкотриенов.
Цитотоксическая, или литическая функция. В конечной стадии активации системы комплемента образуется мембраноатакующий комплекс (МАК) из поздних компонентов комплемента, который атакует мембрану бактериальной или любой другой клетки и разрушает ее.
Фактор С3е, образующийся при расщеплении фактора С3b, обладает способностью вызывать миграцию нейтрофилов из костного мозга, и в таком случае быть причиной лейкоцитоза.
Активация системы комплемента
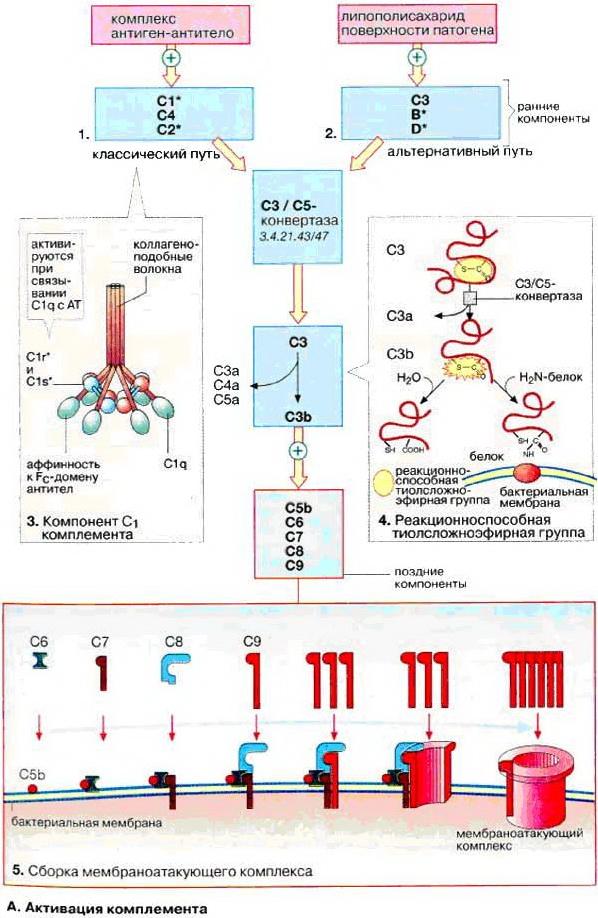
Классический путь
Классический путь запускается активацией комплекса С1 (он включает одну молекулу С1q и по две молекулы С1r и С1s). Комплекс С1 связывается с помощью С1q с иммуноглобулинами классов М и G, связанными с антигенами. Гексамерный C1q по форме напоминает букет нераскрытых тюльпанов, «бутоны» которого могут связываться с Fc участком антител. Для инициации этого пути достаточно единственной молекулы IgM, активация молекулами IgG менее эффективна и требует больше молекул IgG.
С1q связывается прямо с поверхностью патогена, это ведет к конформационным изменениям молекулы С1q, и вызывает активацию двух молекул сериновых протеаз С1r. Они расщепляют С1s (тоже сериновую протеазу). Потом комплекс С1 связывается с С4 и С2 и затем расщепляет их, образуя С2а и С4b. С4b и С2а связываются друг с другом на поверхности патогена, и образуют С3-конвертазу классического пути, С4b2а. Появление С3-конвертазы приводит к расщеплению С3 на С3а и С3b. С3b образует вместе с С2а и С4b С5-конвертазу классического пути. С5 расщепляется на C5a и C5b.C5b остается на мембране и соединяется с комплексом C4b2a3b.Потом соединяются С6, С7, С8 и С9,которая полимеризуется и возникает трубочка внутри мембраны. Тем самым нарушается осмотический баланс и в результате тургора бактерия лопается. Классический путь действует более точно, поскольку так уничтожается любая чужеродная клетка.
Альтернативный путь
Альтернативный путь запускается гидролизом C3 прямо на поверхности патогена. В альтернативном пути участвуют факторы В и D. С их помощью происходит образование фермента СЗbВb. Стабилизирует его и обеспечивает его длительное функционирование белок P. Далее РС3bВb активирует С3, в результате образуется С5-конвертаза и запускается образование мембраноатакующего комплекса. Дальнейшая активация терминальных компонентов комплемента происходит так же, как и по классическому пути активации комплемента. В жидкости в комплексе CЗbВb В заменяется Н фактором и под воздействием дезактивирующего соединения(Н) превращается в С3bi.Когда микробы попадают в организм комплекс СЗbВb начинает накапливаться на мембране. Он соединяется с С5, который расщепляется на C5a и C5b. C5b остается на мембране. Потом соединяются С6, С7, С8 и С9.После соединения С9 с С8, происходит полимеризация С9 (до 18 молекул сшиваются друг с другом) и образуется трубочка, которая пронизывает мембрану бактерии, начинается закачка воды и бактерия лопается.
Альтернативный путь отличается от классического следующим: при активации системы комплемента не нужно образование иммунных комплексов, он происходит без участия первых компонентов комплемента — С1, С2, С4. Он также отличается тем, что срабатывает сразу же после появления антигенов — его активаторами могут быть бактериальные полисахариды и липополисахариды(являются митогенами), вирусные частицы, опухолевые клетки.
Лектиновый (маннозный) путь активации системы комплемента
Лектиновый путь гомологичен классическому пути активации системы комплемента. Он использует лектин, связывающий маннозу, (MBL) — белок, подобный C1q классического пути активации, который связывается с маннозными остатками и другими сахарами на мембране, что позволяет распознавать разнообразные болезнетворные микроорганизмы. MBL — сывороточный белок, принадлежащий к группе белков коллектинов, который синтезируется преимущественно в печени и может активировать каскад комплемента, непосредственно связываясь с поверхностью патогена.
В сыворотке крови MBL формирует комплекс с MASP-I и MASP-II (Mannan-binding lectin Associated Serine Protease, связывающие MBL сериновые протеазы). MASP-I и MASP-II весьма схожи с C1r и C1s классического пути активации и, возможно, имеют общего эволюционного предшественника. Когда несколько активных центров MBL связываются с определенным образом ориентированными маннозными остатками на фосфолипидном бислое болезнетворного микроорганизма, MASP-I и MASP-II активируются и расщепляют белок C4 на C4a и C4b, а белок С2 на C2a и C2b. Затем C4b и C2a объединяются на поверхности болезнетворного микроорганизма, формируя C3-конвертазу, а C4a и C2b действуют как хемоаттрактанты для клеток иммунной системы.
Регуляция системы комплемента
Система комплемента может быть очень опасной для тканей хозяина, поэтому ее активация должна хорошо регулироваться. Большинство компонентов активны только в составе комплекса, при этом их активные формы способны существовать очень короткое время. Если в течение этого времени они не встретятся со следующим компонентом комплекса, то активные формы теряют связь с комплексом и становятся неактивными. Если концентрация какого-то из компонентов ниже пороговой (критической), то работа системы комплемента не приведет к физиологическим последствиям. Система комплемента регулируется специальными белками, которые находятся в плазме крови даже в большей концентрации, чем сами белки системы комплемента. Эти же белки представлены на мембранах собственных клеток организма, предохраняя их от атаки со стороны белков системы комплемента.
Регуляторные механизмы в основном действуют в трех точках.
-
С1. Ингибитор С1 контролирует классический и лектиновый пути активации. Действует двумя путями: ограничивает действие С4 и С2 с помощью связывания C1r- и С1s-протеаз и подобным образом выключает лектиновый путь, удаляя ферменты MASP из MBP-комплекса.
-
С3-конвертаза. Время жизни С3-конвертазы уменьшают факторы ускорения распада. Некоторые из них находятся на поверхности собственных клеток (например, DAF и CR1). Они действуют на С3-конвертазы и классического, и альтернативного путей активации. DAF ускоряет распад С3-конвертазы альтернативного пути. СR1 (C3b/C4b receptor) расположен главным образом на поверхности эритроцитов и отвечает за удаление из плазмы крови опсонизированных иммунных комплексов. Другие регуляторные белки производятся печенью и в неактивном состоянии растворены в плазме крови. Фактор I — сериновая протеаза, расщепляющая C3b и C4b. С4-связывающий белок (C4BP) расщепляет С4 и помогает фактору I расщеплять C4b.Фактор H связывается с гликозаминогликанами, которые есть на собственных клетках, но не на клетках патогенов. Этот белок является кофактором фактора I, а также ингибирует активность C3bBb.
-
С9. CD59 и Гомологичный Фактор Ограничения ингибируют полимеризацию С9 во время образования мембраноатакующего комплекса, не давая ему сформироваться.
Роль системы комплемента при болезнях
Система комплемента играет большую роль во многих болезнях, связанных с иммунитетом.
При болезнях иммунных комплексов комплемент провоцирует воспаление главным образом двумя путями:
-
c C3b и C4b, фиксированными на иммунных комплексах, связываются лейкоцитами, активируемые и привлекаемые в места отложения этих комплексов образовавшимися здесь анафилатоксинами. Так начинается повреждение тканей при синдроме Гудпасчера (системный капиллярит с преимущественным поражением легких и почек по типу гемморагических пневмонита и гломерулонефрита). Для подавления воспалительной реакции на экспериментальных моделях этого заболевания достаточно уменьшить содержание в крови комплемента или нейтрофилов.
-
МАК (мембраноатакующий комплекс), внедряясь в мембрану собственных клеток организма, повреждает мембрану. При этом происходит высвобождение метаболитов арахидоновой кислоты — простагландинов. Этим обусловлено повреждение тканей при мембранозном нефрите, который в эксперименте удается вызвать антителами к субэпителиальным антигенам. Воспалительную реакцию в этом случае не подавляет устранение нейтрофилов, однако она полностью отсутствует у животных, дефицитных по C5.
Белки острой фазы воспаления
Развитие острой системной воспалительной реакции сопровождается повышенной продукцией в печени особых белков, получивших название «белков острой фазы воспаления». Их активная продукция наблюдается через несколько часов с начала воспаления. Основными индукторами их продукции являются провоспалительные цитокины.
С-реактивный белок (СРБ). Является важным фактором регуляции воспалительных процессов и антимикробной защиты организма. СРБ способен связываться с большим числом микроорганизмов и макромолекул. Связывание СРБ с бактериями приводит к разбуханию их капсулы и агглютинации микробов, к фиксации и активации комплемента.
Связывание СРБ с детритом клеток активирует фагоцитоз. Введение СРБ животным увеличивает их выживаемость при экспериментальном микробном заражении и повышает их противоопухолевую резистентность. Взаимодействие СРБ с иммунокомпетентными клетками способно приводить к изменению их метаболизма и функциональной активности. Так, в нейтрофилах и моноцитах наблюдается активация синтеза РНК, белка, секреции ИЛ-1, ИЛ-6, ФНО. СРБ активизирует адгезивные и миграционные свойства лейкоцитов. Способен вызывать агрегацию тромбоцитов. Обнаружена способность СРБ связывать ИЛ-4, и через нейтрализацию этого цитокина переключать гуморальный тип иммунного ответа на клеточный. Показано также способность СРБ связываться с трансформирующим фактором роста β (ТФРβ). Все эффекты, опосредуемые СРБ, неспецифичны.
Сывороточный амилоидный А компонент (СААК). Макромолекулы, характеризующиеся высокой гетерогенностью. Полагают, что СААК участвуют в элиминации липидов микробного происхождения и токсинов, комплексированных с липопротеинами.
α1-Антихимотрипсин. Является ингибитором ряда протеаз (коллагеназы, катепсина, химазы, эластазы), продуцируемых лейкоцитами.
α1-Антитрипсин (α1-антипротеазный ингибитор, α1-АПИ). Составляет 90% общей антипротеолитической активности плазмы. α1-АПИ подавляет активность химотрипсина, трипсина, катепсина, эластазы, нейтральной протеазы, коллагеназы, урокиназы, протеаз комплемента, плазмина, ренина, гиалуронидазы, в некоторой степени – калликреина плазмы, тромбина, а также многих других бактериальных и гранулоцитарных протеиназ. Около 55% α1-АПИ находится вне сосудов, что указывает на его участие в контроле за активностью тканевых протеаз.
α1-АПИ является важным регулятором и контролером активности эластазы, коллагеназы в месте воспаления, выход которых из-под контроля способно привести к деструкции окружающих тканей.
Фибриноген. Относится к классу β-глобулинов. Наиболее значимой функцией фибриногена является участие в формировании тромба и остановке кровотечения. Под влиянием тромбина он превращается в фибрин. Повышенная концентрация фибриногена и фибрина в поврежденной ткани усиливает миграцию в нее гранулоцитов. В интерстициальной ткани фибриноген формирует основу для роста фибробластов и гистиоцитов, что важно для восстановления поврежденной ткани. Продукты деградации фибриногена и фибрина обладают противосвертывающей активностью, способны подавлять процесс формирования фибрина. Это способствует восстановлению кровотока в поврежденной ткани и усиливает его дренажные функции.