Материал: t1_struktura_immunnoy_sistemy
Иммуная система
Иммунная система человека представлена комплексом лимфомиелоидных органов и лимфоидной ткани, ассоциированной с дыхательной, пищеварительной и мочеполовой системами.
К органам иммунной системы относятся: костный мозг, тимус, селезёнка, лимфатические узлы. В состав иммунной системы, помимо перечисленных органов, также входят миндалины носоглотки, лимфоидные (пейеровы) бляшки кишечника, многочисленные лимфоидные узелки, расположенные в слизистых оболочках желудочно-кишечного тракта, дыхательной трубки, урогенитальных путей, диффузная лимфоидная ткань, а также лимфоидные клетки Lamina propria и межэпителиальные лимфоциты.
Главным элементом иммунной системы являются лимфоидные клетки. Общее число лимфоцитов у человека составляет 1012. Вторым важным элементом иммунной системы являются макрофаги. Кроме этих клеток в защитных реакциях организма участвуют гранулоциты. Лимфоидные клетки и макрофаги объединены понятием иммунокомпетентные клетки.
Иммунной системе, как и другим системам организма, свойственны специфические функции. Основной функцией иммунной системы является надзор за макромолекулярным и клеточным постоянством организма, защита организма от всего чужеродного. Иммунная система также обеспечивает контроль и поддержание общего гомеостаза организма, сохранение его уникальности и индивидуальности. Иммунная система вместе с нервной и эндокринной системами регулируют и контролируют все физиологические реакции организма, тем самым обеспечивая жизнедеятельность и жизнеспособность организма. Иммунокомпетентные клетки являются обязательным элементом воспалительной реакции и во многом определяют характер и ход её течения. Важной функцией иммунокомпетентных клеток является контроль и регуляция процессов репаративной и физиологической регенерации тканей.
Свою основную функцию иммунная система осуществляет через развитие специфических (иммунных) реакций, в основе которых лежит способность распознавания «своего» и «чужого» и последующая элиминация чужеродного. Появляющиеся в результате иммунной реакции специфические антитела составляют основу гуморального иммунитета, а сенсибилизированные лимфоциты являются основными носителями клеточного иммунитета.
Специфичность иммунных реакций проявляется в том, что на антиген А вырабатывается антитела или клетки анти-А, которые ни с каким другим антигеном не взаимодействуют, а на антиген В вырабатываются только антитела или клетки анти-В. Феномен «иммунологической памяти» характеризуется тем, что повторный контакт с антигеном вызывает ускоренное и усиленное развитие иммунного ответа, что обеспечивает более эффективную защиту организма по сравнению с первичной иммунной реакцией. Эта особенность вторичной иммунной реакции лежит в основе смысла вакцинации.
В иммунной системе выделяют Т-звено и В-звено или Т-систему иммунитета и В-систему иммунитета. Основными клетками Т-системы иммунитета являются Т-лимфоциты, основными клетками В-системы иммунитета – В-лимфоциты. К главным структурным образованиям Т-системы иммунитета относятся тимус, Т-зоны селезёнки (периартериальные области) и лимфатических узлов (паракортикальные области); В-системы иммунитета – костный мозг, В-зоны селезёнки (центры размножения) и лимфатических узлов (кортикальная зона). Т-звено иммунной системы ответственно за реакции клеточного типа, В-звено иммунной системы реализует реакции гуморального типа. Т-система контролирует и регулирует работу В-системы. В свою очередь, В-система способна оказывать влияние на работу Т-системы. Наиболее ярким доказательством реальности существования двух систем иммунитета у человека являются врождённые иммунологические дефекты. Имеются формы дефектов, когда отсутствует способность вырабатывать антитела при сохранении нормального ответа по Т-клеточному типу. И наоборот, описаны дефекты Т-системы при сохранении относительной активности В-системы иммунитета.
Среди органов иммунной системы различают центральные органы и периферические органы. К центральным органам относятся костный мозг и тимус, к периферическим – селезёнка и лимфатические узлы. В костном мозге из стволовой лимфоидной клетки происходит развитие В-лимфоцитов, в тимусе из стволовой лимфоидной клетки происходит развитие Т-лимфоцитов. По мере созревания Т- и В-лимфоциты покидают костный мозг и тимус и заселяют периферические лимфоидные органы, расселяясь соответственно в Т- и В-зонах.
В центральных органах происходит формирование антигенреактивных Т- и В-лимфоцитов, способных к развитию иммунных реакций. Этот этап развития лимфоцитов протекает без прямого воздействия внешних антигенов. Развитие лимфоцитов в периферических органах происходит под непосредственным воздействием чужеродных антигенов. Этот этап развития лимфоцитов является антиген-зависимым. На этом этапе из антигенреактивных наивных Т-лимфоцитов формируются эффекторные Т-лимфоциты, а из В-лимфоцитов – плазматические клетки, продуценты антител.
Костный мозг
Основой функцией костного мозга является продукция клеток крови и лимфоцитов. Костномозговая ткань пронизана многочисленными гемокапиллярами, которые имеют синусоидный тип. Через эти капилляры происходит миграция зрелых клеток из костного мозга в кровь. Барьерная функция костного мозга в норме обеспечивает выхождение в периферическую кровь только зрелых элементов.
Тимус (вилочковая железа)
Тимус располагается за грудиной. Наибольший его размер относительно тела наблюдается у плода и 1-2-летних детей. До половой зрелости размеры тимуса продолжают увеличиваться. Далее начинается медленная инволюция железы. Однако тимус сохраняется и функционирует в течение всей жизни.
Ежедневно в тимусе образуется около 30 × 107 – 47 × 107 тимоцитов, а покидает орган около 5% клеток. Гемо-тимусный барьер обеспечивает антигеннезависимое развитие Т-лимфоцитов.
Селезенка
Селезенка участвует в следующих процессах: обеспечивает иммунные реакции организма, в ней происходит продукция лимфоцитов в ответ на антигенный стимул, обеспечивает отбор и элиминацию функционально неактивных эритроцитов и лейкоцитов, кровяных пластинок, служит депо крови.
Лимфатический узел
Лимфоидные узелки и мозговые тяжи являются В-зонами лимфатических узлов. Здесь происходит размножение и превращение В-лимфоцитов в плазмоциты.
На границе между корковым и мозговым веществом располагается паракортикальная зона. Она содержит, главным образом, Т-лимфоциты. В этой зоне происходит размножение и дифференцировка Т-лимфоцитов в эффекторные клетки.
Развитие Т-лимфоцитов в тимусе
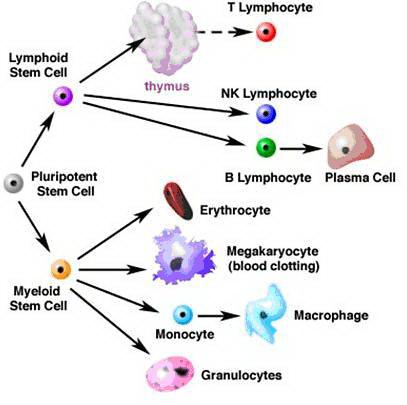
Развитие Т-клеток в тимусе происходит под непосредственным влиянием и в результате прямых контактов тимоцитов со стромальными эпителиоцитами, клетками-кормилицами, макрофагами тимуса, а также под влиянием гормонов тимуса. В тимусе Т-клетки в процессе своего развития приобретают способность распознавать антиген в контексте с молекулами ГКГ и толерантность к собственным тканевым антигенам.
Маркеры, появляющиеся в процессе дифференцировки лимфоцитов, получили название дифференцировочных маркеров (CD) (cluster of differentiation) или дифференцировочных антигенов.
В процессе развития в тимусе происходит элиминация Т-клеток, способных взаимодействовать с АГ собственных тканей. В данном случае включаются механизмы отрицательной селекции клеток. Тимоциты, несущие рецепторы, способные распознавать собственные антигены в комплексе с молекулами ГКГ, в результате взаимодействия с ними подвергаются апоптозу, что приводит к элиминации соответствующего клона аутореактивных Т-лимфоцитов.
Ежедневно в тимусе вырабатывается порядка 30 × 107 – 47 × 107 клеток. Большая часть из них (95%) гибнет в органе, и только небольшое количество клеток продолжает своё дальнейшее развитие.
Клетки, прошедшие полную программу развития в тимусе и выдержавшие условия отбора на специфичность, покидают орган и мигрируют в периферическую лимфоидную ткань.
В итоге, в тимусе продуцируются две категории наивных антигенреактивных клеток: Т-лимфоциты воспаления/хелперы (CD4+, Т H0) (которые на периферии под влиянием АГ трансформируются в Т-лимфоциты хелперы гуморального иммунитета (Т H2) и Т-хелперы клеточного иммунитета (Т H1) и Т-цитотоксические лимфоциты (CD8+ ).
Миграция Т-лимфоцитов на периферию
Процесс миграции тимуспроизводных клеток в периферические лимфоидные органы контролируется адгезивными молекулами, которые находятся на поверхности как Т-лимфоцитов, так и на клетках тех органов, которые они заселяют.
Развитие В-лимфоцитов в костном мозге
Развитие В-лимфоцитов в течение всего постэмбрионального периода протекает в костном мозге. Под воздействием клеточного костномозгового микроокружения и гуморальных факторов костного мозга из стволовой лимфоидной клетки формируются В-лимфоциты
На определенной стадии развития В-лимфоцитов происходит экспрессия поверхностных мономерных иммуноглобулинов М. Эти структуры и являются антиген-распознающими рецепторами В-клеток. Антигенная специфичность рецепторов генетически детерминирована. На последующем этапе развития В-лимфоцитов происходит ориентация клеток на синтез антител определенного класса. Появляются В-лимфоциты, которые экспрессируют наряду с IgМ молекулы класса IgA или IgG.
Далее происходит экспрессия на клетках IgD. С экспрессией на лимфоцитах иммуноглобулинов D завершается этап антиген-независимого созревания В-клеток. Таким образом, на зрелых В-лимфоцитах поверхностные Ig-молекулы могут быть представлены следующими классами: 1) IgM, IgD; 2) IgM, IgA, IgD; 3) IgM, IgG, IgD. При этом все иммуноглобулины, представленные на одной В-клетке, имеют одинаковый идиотип, так как кодируются одними и теми же генами VH и VL.
Зрелые В-лимфоциты характеризуются наличием поверхностного IgD, высокой плотностью рецепторов к СЗ компоненту комплемента и Fc- фрагменту Ig, способностью трансформироваться в бластные формы под влиянием В-митогенов (ЛПС, PWM) и способностью трансформироваться под влиянием антигенов в антителообразующие клетки.
Строение в-клеточного рецептора
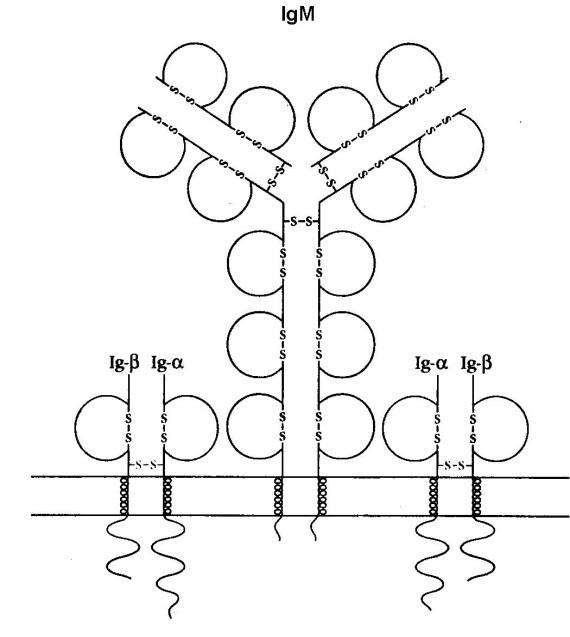
Антигенсвязывающей молекулой В-лимфоцитов является мономерный иммуноглобулин М, связанный с мембраной клетки. Молекула IgМ также плотно связана с гетеродимерами Ig-a/Ig-b. Этот комплекс молекул и формирует единый В-клеточный рецептор (ВКР). Каждый гетеродимер содержит экстрацеллюлярный Ig-подобный сегмент, мембранный участок и цитоплазматический хвост, который связан с внутриклеточными сигнальными молекулами. Гетеродимеры Ig-α /Ig-β участвуют в трансдукции сигнала с IgМ в ядро клетки. Дефектность молекул Ig-α /Ig-β нарушает проведение сигнала с рецептора внутрь клетки.
В костном мозге в процессе развития В-лимфоцитов происходит селекция клонов, способных реагировать только на чужеродные антигены. Клетки, экспрессирующие иммуноглобулиновые рецепторы к своим собственным антигенам, либо погибают в результате запуска механизма апоптоза, либо переходят в состояние ареактивности (анергии). Апоптоз развивается в тех случаях, когда распознавание антигена как «своего» происходит на поверхности клетки. Распознавание свободного антигена приводит к анергии.
В результате реализации механизма отрицательной селекции в В-клеточной популяции формируется толерантность к собственным антигенам.
В-клетки, прошедшие полную программу развития в костном мозге, покидают его и мигрируют на периферию, где заселяют B-зоны лимфоидных органов и лимфоидную ткань, ассоциированную со слизистыми покровами. Хоминг лимфоцитов к соответствующим зонам определяется их поверхностными адгезивными молекулами (селектинами, интегринами, адгезинами суперсемейства иммуноглобулинов, муциноподобными молекулами).
Кластеры дифференцировки
Кластер дифференцировки (англ. cluster of differentiation, cluster designation; сокращённо CD) — номенклатура дифференцировочных антигенов лейкоцитов человека. Данная классификация была предложена в 1982 году для идентификации и исследования поверхностных мембранных белков лейкоцитов. CD-антигенами (или иначе CD-маркёрами) могут быть белки, которые служат рецепторами или лигандами, участвующими во взаимодействии клеток между собой и являющимися компонентами каскада определённых сигнальных путей. Однако, они могут быть и белками, выполняющими другие функции (например, белки клеточной адгезии). Список CD-антигенов, внесённых в номенклатуру, постоянно пополняется и в настоящее время содержит 350 CD-антигенов и их подтипов.
Иммунофенотипирование
Система кластеров дифференцировки применяется в иммунофенотипировании для отнесения клеток к тому или иному типу по представленным на клеточных мембранах молекулам-маркёрам. Наличие определённых молекул может быть ассоциировано с соответствующими иммунными функциями. Хотя наличие одного типаCD обычно не позволяет точно определить популяцию клетки (за исключением нескольких примеров), сочетания маркёров позволяют определить её достаточно чётко.
СD молекулы используемые для сортировки клеток в различных методах таких как проточная цитометрия.
|
Тип (популяция) клеток |
CD маркеры |
|
Стволовые клетки |
CD34+, CD31- |
|
Все лейкоциты |
CD45+ |
|
Гранулоциты |
CD45+, CD15+ |
|
Моноциты |
CD45+, CD14+ |
|
T-лимфоциты |
CD45+, CD3+ |
|
Т-хелперы |
CD45+, CD3+, CD4+ |
|
Цитотоксические Т-лимфоциты |
CD45+, CD3+, CD8+ |
|
B-лимфоциты |
CD45+, CD19+ или CD45+, CD20+ |
|
Тромбоциты |
CD45+, CD61+ |
|
Естественные киллеры |
CD16+, CD56+, CD3- |
Два наиболее широко используемых CD маркера — CD4 и CD8, которые соответственно являются характерными для T-хелперов и цитотоксических Т-лимфоцитов. Эти молекулы определяются в сочетании с CD3+, так и с другими маркёрами для других популяций клеток (некоторые макрофаги экспрессируют низкие уровни CD4; дендритные клетки имеют высокие уровни CD8). Вирус иммунодефицита человека (ВИЧ) связывает CD4 и хемокиновый рецептор на поверхности T-хелперов для проникновения в клетку. Таким образом, количество CD4 и CD8 T-лимфоцитов в крови часто используется для мониторинга развития ВИЧ инфекции.
Т-лимфоциты
Т-лимфоциты. Интактные Т-лимфоциты крови и лимфоидных органов имеют морфологию малых лимфоцитов. Т-лимфоциты выполняют следующие функции:
– являются основными эффекторами клеточного иммунитета (эти клетки опосредуют клеточные цитотоксические реакции, а также реакции ГЗТ);
– являются регуляторами воспаления, иммунных реакций и гемопоэза;
– участвуют в процессах репаративной и физиологической регенерации различных тканей.
Среди Т-лимфоцитов различают две фенотипические субпопуляции клеток – CD4+-клeтки и СD8+-клетки. По функциональным характеристикам в популяции Т-лимфоцитов выделяют Т-хелперы гуморального иммунитета, Т-хелперы клеточного иммунитета, Т-супрессоры, Т-цитотоксические клетки. Т-хелперы гуморального и клеточного иммунитета имеют единого предшественника – ТH0-клетки, из которых они генерируются в ходе иммунного ответа.
Т-лимфоциты хелперы гуморального иммунитета (Т H2,CD4+). Клетки несут фенотипический маркер CD4, характеризуются способностью продуцировать интерлейкины 4, 5, 6. Т-хелперы участвуют в качестве вспомогательных клеток в индукции гуморального иммунитета, развитии аллергических реакций, в контроле и регуляции дифференцировки гемопоэтических стволовых клеток.
Т-лимфоциты хелперы клеточного иммунитета (Тн1, CD4+, ).Клетки характеризуются поверхностным маркером CD4 и способностью продуцировать интерлейкин-2, ИНФγ, ФНОβ и ГМ-КСФ. Участвуют в качестве вспомогательных клеток в развитии клеточного иммунитета. Клетки участвуют в индукции воспаления, активации антибактериальных свойств макрофагов, реакции ГЗТ, активации фибробластов соединительной ткани и синтезе в них коллагена.
Т-лимфоциты супрессоры. Т-лимфоциты супрессоры принимают участие в контроле и ограничении развития гуморальных и клеточных иммунных реакций, способствуют их окончанию, поддерживают толерантность к собственным антигенам, блокируют развитие аутоиммунных реакций.
Среди Т-лимфоцитов супрессоров различают:
1) антигенспецифические Т-супрессоры;
2) неспецифические Т-супрессоры.
Следует заметить, что в настоящее время имеются серьезные сомнения в существовании отдельной линии Т-клеток, обладающей только супрессорными свойствами. В ряде работ показано, что супрессивное действие на развитие реакций как гуморального, так и клеточного типов способны оказывать как CD4+, так и CD8+, – клетки.
Т-цитотоксические лимфоциты (ЦТЛ) (CD8+). Клетки несут фенотипический маркер CD8. Из наивных ТЦЛ в ходе развития клеточной иммунной реакции генерируются Т-киллеры, способные оказывать прямое цитотоксическое действие на чужеродные клетки, измененные свои клетки и клетки, инфицированные вирусами.
B-лимфоциты
У человека интактные B-лимфоциты крови и лимфоидных органов имеют морфологию малых лимфоцитов. Основной функцией B-лимфоцитов является выработка антител. В-лимфоциты, в зависимости от экспрессии на своей поверхности отдельных классов иммуноглобулинов, делятся на следующие субпопуляции:
1) B-лимфоциты IgM+, IgD+;
2) В-лимфоциты IgM+, IgA+, IgD+;
3) B-лимфоциты IgM+, IgG+, IgD+.
Экспрессия всеми В-клетками мембранных иммуноглобулинов позволяет осуществляться клональному отбору под действием антигена. При созревании, стимулировании антигеном и пролиферации существенно меняется набор маркеров В-клеток. По мере созревания В-клетки переключаются от синтеза IgM и IgD на синтез IgG, IgA, IgE (при этом у клеток сохраняется способность синтезировать также IgM и IgD — вплоть до трёх классов одновременно). При переключении синтеза изотипов антигенная специфичность антител сохраняется. Различают следующие типы зрелых В-лимфоцитов: