Материал: Роль полушарий головного мозга в восприятии зрительной информации
мозг сетчатка затылочный зрительный
ВОСПРИЯТИЕ ЗРИТЕЛЬНОЙ ИНФОРМАЦИИ КОРОЙ
ГОЛОВНОГО МОЗГА
2.1 Исследования
зрительной коры головного мозга
Первичную зрительную кору называют также «полем 17» или «стриарной корой». Она состоит из весьма упорядоченных слоев и представляет собой структуру, уникальную по своей сложности во всей нервной системе. Для всей коры большого мозга характерна слоистая структура, состоящая, как правило, из шести слоев - от I до VI, начиная с внешней поверхности. Слои различаются по количеству содержащихся в них нейронов. Однако в зрительной коре человека и обезьян эти слои в свою очередь подразделяются, что особенно характерно для слоев IV и V. У приматов можно выявить более 12 слоев зрительной коры [9, 511 c.].
Как показывает схема, разработанная Е. Г.
Школьник-Яррос (Рис. 2.1), большинство идущих от сетчатки волокон, прерываясь в
ядрах наружного коленчатого тела, направляется к той части затылочной коры,
которая располагается в полюсе затылочной области и затем идет по ее медиальной
поверхности. Эта часть (поле 17-е Бродмана) с полным основанием может быть
оценена как первичное поле коркового ядра зрительного анализатора. Некоторое
количество афферентных волокон, прерываясь в наружном коленчатом теле и
поступая затем во «вторичные» (вентральное и заднее) ядра зрительного бугра,
доходит до тех отделов коры затылочной области (поле 18-е и 19-е Бродмана),
которые могут быть обозначены как «широкая зрительная сфера» (Петцль, 1928)
коркового ядра зрительного анализатора.
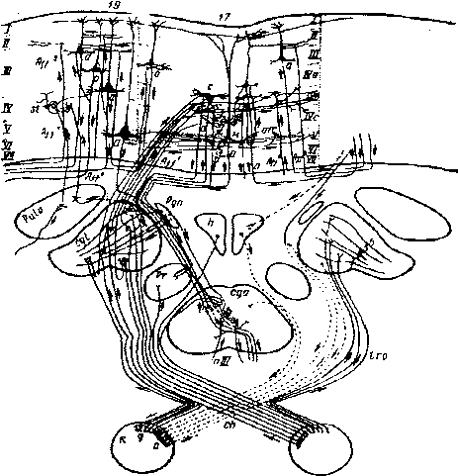
Рис. 2.1 Схема связей и переключений зрительного анализатора (по Е. Г. Школьник-Яррос): Вверху - схематическое изображение полей 17-го и 19-го. Римcкими цифрами обозначены слои коры: слева-для поля 19-го, справа -для поля 17-го. Поле 18-е, располагающееся между полями 17-м и 19-м, на схеме не обозначено. В средней части схемы даны подкорковые образования, внизу - сетчатка глаз. Показаны центростремительные (сплошные линии) и центробежные (прерывистые линии) связи сетчатки. Pulv - подушка зрительного бугра; Cge- наружное коленчатое тело; pgn- прегеникулярное ядро; h - гипоталамус; рг - претектальное ядро; cqa - переднее двухолмие; - глазодвигательный нерв; ch-хиазма; R - сетчатка; g- ганглиозные клетки сетчатки; а - амакринные клетки сетчатки; tro - зрительный тракт. Стрелками обозначено направление передачи возбуждения. Для составления схемы использованы собственные и литературные данные (Минковский, Ле Гро Кларк, Поляк, Кахал, Новохатский, Чоу и др.)
Из всех перечисленных полей зрительной коры берут начало как афферентные волокна, направляющие к подкорковым зрительным ядрам и от них к сетчатке, так и ассоциативные волокна, идущие от 17-го поля к 18-му и 19-му и обратно. Все вместе они составляют тот рефлекторный аппарат зрительной коры, который осуществляет сложные акты зрительного восприятия.
Столь же существенным, как и фугально-петальное строение проводящего аппарата зрительной рецепции, является и топическая организация самой зрительной коры.
Еще со времени Брауэра (1917-1932, сводное изложение 1936) хорошо известно, что волокна, идущие от сетчатки, располагаются в строгом порядке, так что отдельные части наружного коленчатого тела и проекционных отделов зрительной коры соответствуют определенным частям сетчатки. В силу этого раздражение соответствующих точек зрительной коры вызывает фотопсии в определенных частях зрительного поля, а разрушение этих участков зрительной коры приводит к строго определенному выпадению соответствующих (контралатеральных) участков зрительного поля, известных под названием гемианопсий или скотом. Тщательное исследование этих явлений было в последнее время проведено Тейбером (1960).
Таким образом, и эти зоны (аналогично 22-му полю височной коры) относятся к тем разделам, которые приобрели название «вторичных» корковых зон [7, 111-113 c.].
Проекции изображений видимого мира от каждого из латеральных коленчатых ядер передаются по волокнам так называемой зрительной радиации в правую и левую части первичной зрительной коры. Однако эти проекции на корковом уровне уже не представляют собой точных отображений внешнего мира. Область коры, получающая информацию от центральной ямки - зоны наивысшей остроты зрения, примерно в 35 раз больше участка, отображающего кружочек той же величины на периферии сетчатки. Таким образом, информация, идущая от центральной ямки, имеет для коры неизмеримо большее значение, чем информация от других частей сетчатки [18, 142 c.].
Нейронографические исследования, проведенные Бонином, Гэрол и Мак-Кэллоком (1942) и др., показали, что этому архитектоническому строению вторичных полей затылочной области соответствуют определенные физиологические свойства. Если верить данным указанных авторов, местное раздражение 19-го поля вызывает столь же широкий, но обратный по знаку тормозящий эффект [7, 114 c.].
Исследования, начатые Хоффом и Петцлем (1930) и широко продолженные Пенфилдом (1941, 19.54 и др.), показали, что раздражение проекционных отделов затылочной области приводит лишь к возникновению элементарных зрительных ощущений (цветные круги, туман, языки пламени и т. п.), в то время как раздражение вторичных отделов зрительной коры вызывает более сложные, содержательные, зрительные галлюцинации с оживлением зрительных образов, сложившихся у больного в его предшествующем опыте. Различный по своей сложности характер работы отдельных участков зрительной коры получает в этих опытах свое фактическое подтверждение [7, 114 c.].
Следует отметить, что площадь вторичных зон затылочных отделов мозговой коры по сравнению с ее первичными зонами существенно увеличивается в процессе эволюции: если у низших обезьян (с их мощно развитой зрительной корой) площадь первичной коры значительно преобладает над площадью вторичной, то у человека это отношение является обратным: площадь вторичной зрительной зоны (18-е поле Бродмана) явно преобладает над площадью первичной зрительной коры (17-е поле Бродмана).
Данные, отчетливо показывают, что роль аппаратов, связанных с синтезом и переработкой получаемой зрительной информации, значительно возрастает у человека по сравнению с его предшественниками на эволюционной лестнице.
Как показали морфологические исследования, общее число нервных клеток в отдельных слоях первичной и вторичной коры затылочной области различно. Если в первичной зоне зрительной коры (17-е поле) преобладают клетки, расположенные в IV (афферентном) слое, то во вторичной зоне зрительной коры (18-е поле) преобладают клетки, расположенные в верхних (II и III) «ассоциативных» слоях коры.
Эти цифры отчетливо указывают на функциональное различие первичных и вторичных полей затылочной области. Этот факт подтверждается и физиологическими исследованиями, проведенными на нейронном уровне. Как показывают нейронографические исследования (Бонин, Гэроль, Мак-Кэллок, 1942), раздражение вторичных отделов зрительной коры распространяется на более обширные области, чем раздражение ее первичных отделов, переходя иногда даже на соответствующие области противоположного полушария [20, 105 c.].
Описанные морфологические и физиологические особенности строения вторичных отделов зрительной коры определяют ту роль, которую она играет в организации сложнейших процессов зрительного восприятия, с особенной отчетливостью выступающую в опытах, проведенных выдающимися неврологами и нейрохирургами (Петцлем, Ферстером, Пенфилдом) во время операций на этих отделах мозговой коры. Они показали, что раздражение первичных (проекционных) отделов затылочной области слабым электрическим током вызывает у больного появление элементарных зрительных ощущений в виде фотопсий (светящихся точек, языков пламени, цветовых пятен и т.п.). Характерно, что эти явления возникают в строго определенных участках зрительного поля: раздражение правой затылочной области ведет к возникновению фотопсий в левой, а раздражение левой затылочной области - к появлению фотопсий в правой части зрительного поля.
Совершенно иные явления возникают при раздражении вторичных отделов зрительной коры. В этих случаях электрическое раздражение того или иного пункта коры вызывает не элементарные зрительные ощущения, а сложные оформленные зрительные галлюцинации (образы цветов, животных, знакомых лиц и т.д.). Иногда всплывают целые сложные сцены, например, больной видит своего знакомого, идущего со стороны и делающего ему знак рукой, и т. п. Следует отметить, что эти галлюцинации не привязаны к определенному участку зрительного поля и носят не топический, а смысловой характер.
Естественно, что эти галлюцинации отражают прежний зрительный опыт субъекта, и, следовательно, раздражение вторичных отделов зрительной коры пробуждает следы тех зрительных образов, которые хранятся в долговременной зрительной памяти человека.
Таким образом, вторичные отделы зрительной коры, обеспечивая сложное и распространенное протекание возбуждений, играют роль аппарата, синтезирующего зрительные раздражения и организующего их в определенные системы. Эти отделы коры играют решающую роль в обеспечении более высокого уровня переработки и хранения зрительной информации.
Такой характер работы вторичных зон затылочной коры - обеспечение синтеза зрительных возбуждений и создание тем самым физиологической основы для сложного зрительного восприятия - становится совершенно очевидным из наблюдений над изменениями зрительных процессов, возникающими при локальных поражениях вторичных областей зрительной коры.
Как показывают клинические наблюдения, локальные поражения этих областей не приводят к гемианопсиям, не вызывают выпадения отдельных участков зрительного поля, не ведут к снижению остроты зрения. Важным симптомом поражения этих областей является нарушение интегрального восприятия целых зрительных комплексов, невозможность объединять отдельные впечатления в целостные образы, что приводит к возникновению феномена не узнавания реальных предметов и их изображений [4, 143-145 c.].
Как уже было указано ранее, первичные зоны затылочной коры являются тем местом, где кончаются волокна, идущие от сетчатки глаз; эти волокна идут сначала в составе зрительного нерва, затем не полностью перекрещиваются в хиазме, продолжая свой путь в зрительном тракте, причем зрительный тракт правого полушария включает волокна, передающие возбуждения, которые воспринимаются левыми половинами зрительного поля обоих глаз, а зрительный тракт левого полушария - волокна, несущие возбуждения, воспринимаемые правыми половинами зрительного поля обоих глаз; волокна зрительного тракта оканчиваются в наружном коленчатом теле, где начинается новый зрительный путь, который веером располагается внутри височной области (зрительное сияние) и заканчивается в первичном (проекционном) поле затылочной коры.
Все приведенные факты показывают, что процесс зрительного восприятия является сложной функциональной системой, опирающейся на совместную работу целого комплекса корковых зон, и что каждая из этих зон вносит свой собственный вклад в построение активной перцепторной деятельности.
Изучая таким образом связи между слоями и
зонами, исследователи выявили по меньшей мере еще пять уровней интеграции
зрительной информации в коре. «Наивысшим» из них оказался уровень, связанный со
зрительными полями лобной коры. Они примыкают к так называемой ассоциативной
коре, где происходит объединение различных видов сенсорной информации.
Возможно, что эта корковая зона имеет прямые связи и с лимбической системой [4,
148-149 c.].
2.2 Нарушение
зрительных функций при локальных поражениях мозга
Чуть больше 150 лет назад знаменитый французский антрополог и анатом Поль Брока сообщил, что при вскрытии двух больных, страдавших расстройством речи, он обнаружил поражение одной и той же области левого полушария - заднелобной. После нескольких лет раздумий и наблюдений Брока в статье, опубликованной в шестом томе «Бюллетеня антропологического общества» за 1865 г., заявил: «Мы говорим левым полушарием».
Еще десять лет спустя его соотечественник Клодт Вернике нашел, что при кровоизлияниях в височную область того же полушария больной перестает понимать речь, хотя и может говорить: она превращается для него в бессмысленный шум. «Говорящее» полушарие из уважения к столь важному делу, как речь, назвали доминантным, господствующим, а «безмолвное» - субдоминантным, подчиненным. Терминология способствовала тому, что наибольшее внимание исследователи уделяли доминантному полушарию, и только в самые последние годы выяснили: и субдоминантное достойно самого пристального изучения [10, 63 c.].
Не каждый человек «говорит левым полушарием». Даже если он правша, это будет лишь в девяноста пяти случаях из ста, а у оставшихся пяти доминантным окажется правое. У левшей соотношение тоже не абсолютно: шестьдесят пять из ста подчиняются правилу «доминантное полушарие противоположно ведущей руке», остальные же, хотя и пишут левой рукой, говорят все же «обычным», левым полушарием [10, 65 c.].
Чем дальше исследует наука специализацию полушарий, тем яснее выявляются поразительные разделения функций. Масса данных была получена при исследованиях «душевной слепоты» (по нынешней терминологии - агнозий), то есть особых расстройств мозговой деятельности.
Первым стал заниматься агнозиями английский невропатолог Хьюлинг Джексон, после того как в 1874 г. подметил, что некоторые больные перестают при поражениях правого полушария узнавать лица. В полном порядке сетчатка, здоров зрительный нерв, нет ощущений ни близорукости, ни дальнозоркости, нормально поле зрения, и вдруг человек не может сказать, кому принадлежит лицо, глядящее на него в упор из зеркала. С тех пор описано множество агнозий правого и левого полушарий. Бывает, видит больной предметы, а телефон называет часами, грушу - цветком, садовая скамейка превращается для него в диван. Стрелки же на часах ставит совершенно правильно, именно на то время, которое называет врач. Или, бывает, не способен назвать вещь, пока не пощупает ее. Или видит буквы, но воспринимает их просто как рисунки, хотя сразу вспоминает значение, едва обведет контур пальцем, подобно маленькому ребенку. Или потеряна способность читать, букв больной не узнает, а цифрами оперирует по-прежнему свободно [4, 158 c.].
Нередко агнозии - следствие кровоизлияний или опухолей мозга. Сопоставляя их с результатами операций, ученые судят о том, какие области полушарий какими функциями заведуют [4, 160 c.].
Немало данных было получено, когда здоровые люди решали зрительные задачи отдельно левым или правым полушарием. Ведь правая и левая половины сетчатки каждого глаза соединены соответственно с левым и правым полушариями. Если демонстрировать изображения так быстро, чтобы половинки мозга не успели обменяться информацией, можно быть уверенным: ответ (словом или действием) - следствие работы только одного полушария.
В конце 50-х гг. были опубликованы результаты опытов, показавших огромную роль таких связей и потенциальную возможность независимой работы каждого из полушарий. Речь идет о знаменитых «калифорнийских кошках», названных так потому, что эксперименты над ними велись в Калифорнийском технологическом институте. Там работал нейрофизиолог Р. Сперри, который перерезал нескольким кошкам мозолистое тело. После операции ожидали чего угодно, только не того, что каждая половинка мозга станет работать самостоятельно, как если бы в животном было заключено сразу два живых существа.
Узнали это, обеспечив связь каждого глаза только с одним полушарием. Для этого была перерезана хиазма - перекрест зрительных нервов. Теперь информация от каждой сетчатки (вернее, от соответствующей ее половины) шла только в одноименную зрительную кору. Образовались два комплекса «глаз - полушарие», и экспериментатор принялся обучать их отдельно. Для этого противоположный глаз закрывали повязкой, и, скажем, левый комплекс приучался к тому, что пища лежит за дверцей, на которой нарисован круг, а правый комплекс - что на ней квадрат. В итоге кошка как бы раздваивалась: у нее формировалось два набора условных рефлексов, и любой можно было включить или выключить, просто закрыв ей соответствующий глаз.