Материал: Роль полушарий головного мозга в восприятии зрительной информации
Роль полушарий головного мозга в восприятии зрительной информации
КУРСОВАЯ РАБОТА
Роль
полушарий головного мозга в восприятии зрительной информации
ВВЕДЕНИЕ
По данным некоторых ученых 70% всех сведений человек получает из окружающего мира с помощью зрения, другие полагают, что цифра должна быть увеличена до 90%. Зрительная система реагирует на световые раздражители. Мы видим объекты потому, что они отражают свет. Цвета, которые мы различаем, определяются тем, какую из частей видимого светового спектра отражает или поглощает предмет. Основная функция зрения состоит в различении яркости, цвета, формы, размеров наблюдаемых объектов. Наряду с другими анализаторами зрение играет большую роль в регуляции положения тела и в определении расстояния до объекта.
Изучение работы зрительного анализатора - одна из актуальных задач современной нейрофизиологии. Среди всех анализаторов зрительный отличается не только структурной и функциональной сложностью, но прежде всего долей и разнообразием сенсорной информации, на него приходящейся.
Исследование зрительного анализатора многопланово в любом аспекте, а в нейрофизиологическом - особенно. Из всех видов зрительного анализа (форма, цвет, движение, пространство) для человека ведущим является анализ формы и движения. Сформировавшаяся в процессе эволюции надежная и быстродействующая система зрительного анализа движущихся объектов привлекает особое внимание исследователей, поскольку оказалось возможным создать ряд технических устройств, в том числе и для слежения за движущимся объектом, на основе использования принципов организации нейронных структур этой сенсорной системы
Немецкий физик Герман Гельмгольц, изучавший во второй половине XIX века глаза животных, установил, что зрительная информация отображается на сетчатке точно так же, как и в любой простой камере с линзой: глаз создает перевернутое и уменьшенное изображение предметов. С этих простых сведений началось накопление того богатства знаний о зрительной системе, которым мы сейчас располагаем. Действительно, мы куда лучше понимаем, как реконструируется зрительный образ окружающего нас мира, чем то, как интерпретируется любая другая сенсорная информация.
Объектом курсовой работы является головной мозг.
Предметом - влияние полушарий головного мозга на восприятие зрительной информации.
Целью курсовой работы является изучить механизм восприятия зрительной информации полушариями голоного мозга.
Задачи курсовой работы:
· изучить научную литературу по данной проблеме;
· изучить анатомические структуры зрительной системы;
· исследовать механизм восприятия
зрительной информации полушариями головного мозга.
. СТРОЕНИЕ ЗРИТЕЛЬНОЙ СИСТЕМЫ
Зрительный анализатор представляет собой сложную
многозвеньевую систему (рис. 1.1). Он состоит из периферического отдела -
глаза, промежуточных - подкорковых зрительных центров и конечного звена - зрительного
центра в коре головного мозга. Все уровни зрительной системы соединены друг с
другом проводящими путями [18, 26 c.].
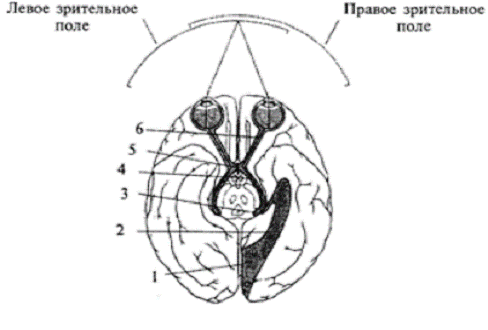
Рис. 1.1 Схема строения зрительной системы
человека: 1 - первичная зрительная кора; 2 - зрительная радиация; 3 -
латеральное коленчатое тело; 4 зрительный тракт; 5 - зрительный перекрест; 6 -
зрительный нерв
1.1 Строение глаза
Глаз позвоночных животных имеет шарообразную (или близкую к таковой) форму (рис. 1.2). Он образован несколькими оболочками. Склера - плотная наружная непрозрачная соединительнотканная оболочка - в передней части глазного яблока переходит в прозрачную роговую оболочку, или роговицу. Под склерой лежит сосудистая оболочка, образованная сетью кровеносных сосудов.
Спереди сосудистая оболочка утолщается и
переходит сначала в ресничное тело и далее - в радужную оболочку, которые
состоят из гладких мышечных волокон, кровеносных сосудов и пигментных клеток.
Мышечные волокна ресничного тела прикреплены к склере. В центре радужной оболочки
расположено отверстие - зрачок. С внутренней стороны сосудистой оболочки
находится слой клеток пигментного эпителия, к нему прилегает самая внутренняя
оболочка глаза - сетчатая оболочка, или ретина, выполняющая основную функцию
глаза - преобразование светового раздражителя в нервное возбуждение и первичную
обработку сигнала. Волокна самой внутренней части сетчатки переходят в
зрительный нерв. Между роговой и радужной оболочками расположена полость,
наполненная жидкостью, - передняя камера глаза. За радужной оболочкой находится
прозрачное тело, имеющее форму двояковыпуклой линзы, - хрусталик, прикрепленный
к ресничному телу. За хрусталиком вся полость глазного яблока заполнена
студенистым содержимым - стекловидным телом.
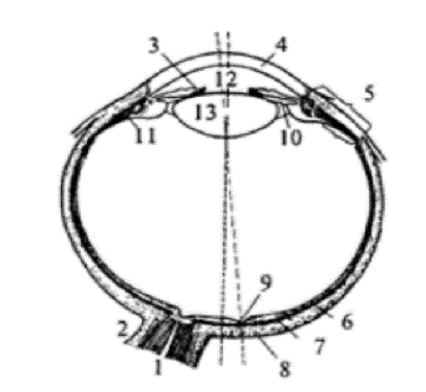
Рис. 1.2 Строение глаза человека: 1 - зрительный
нерв; 2- твердая мозговая оболочка; 3 - радужная оболочка; 4 - роговица; 5 -
цилиарное тело; 6 - сетчатка; 7 - сосудистая оболочка; 8 - склера; 9 -
центральная ямка; 10 - задняя камера; 11 - цилиарная мышца; 12 - передняя
камера; 13 - хрусталик
1.2 Строение
сетчатки
Сетчатка - cветовоспринимающий аппарат глаза - выстилает заднюю и боковые внутренние поверхности глазного яблока (рис. 1.3). Сетчатка состоит из 10 слоев.
· Пигментный эпителий. Самый наружный слой сетчатки содержит плоские эпителиальные клетки, заполненные пигментом. Дает свои отростки во второй слой.
· Слой наружных сегментов фоторецепторов (палочек и колбочек).
· Наружная пограничная мембрана. Образована отростками мюллеровых клеток.
· Наружный зернистый (наружный ядерный) слой. Образован внутренними сегментами и ядерно-плазматическими телами фоторецепторов.
· Наружный сетевидный слой. Содержит синоптические контакты между фоторецепторами, биполярными и горизонтальными клетками.
· Внутренний зернистый (внутренний ядерный) слой. Состоит из тел горизонтальных, биполярных, амакриновых и мюллеровых клеток, а также содержит центробежные волокна сетчатки.
· Внутренний сетевидный слой. Образован синапсами между биполярными, ганглиозными и амакриновыми клетками.
· Слой ганглиозных клеток.
· Слой нервных волокон. Образован аксонами ганглиозных клеток.
· Внутренняя пограничная мембрана.
Образована (аналогично наружной) отростками мюллеровых клеток.
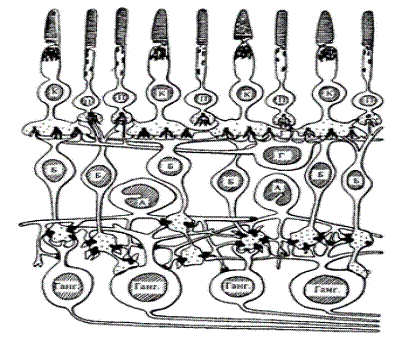
Рис. 1.3 Строение сетчатки: К - колбочки; П -
палочки; Б - биполяры; Г - горизонтальная клетка; А - амакриновая клетка; Ганг.
- ганглиозные клетки
Наружная часть сетчатки (та, что прилегает к сосудистой оболочке и ограничена слоем пигментных клеток) образована специальными светочувствительными клетками - фоторецепторами. У большинства позвоночных фоторецепторы различаются по своей форме и называются палочками и колбочками.
Центральная часть сетчатки представлена
биполярными клетками, имеющими по два относительно длинных отростка, одним из
которых они контактируют с фоторецепторами, другим - с ганглиозными клетками
сетчатки, которые в свою очередь, составляют ее внутреннюю часть. Таким образом
фоторецепторы, биполярные и ганглиозные клетки представляют собой три последовательных
звена переработки зрительной информации. На уровне между рецепторами и
биполярами имеются специализированные клетки с горизонтальным расположением
отростков, которые регулируют передачу возбуждения от рецепторов к биполярам,
они называются горизонтальными. Между биполярами и ганглиозными клетками,
располагаясь как бы симметрично горизонтальным, находятся так называемые
амакриновые клетки, которые «управляют» передачей электрических сигналов от
биполяров к ганглиозным клеткам. Наконец, аксоны ганглиозных клеток формируют
зрительный нерв, который пронизывает сетчатку в противоположном направлении и
входит в полость черепа. В месте вхождения в сетчатку зрительного нерва
фоторецепторы отсутствуют, эта область получила название слепого пятна [12,
304-306 c.].
1.3 Физиология
сетчатки
Общая схема нейрофизиологических процессов, протекающих в сетчатке, выглядит следующим образом:
· Фотохимические превращения в наружном сегменте фоторецептора в результате действия светового стимула приводят к генерации фоторецепторных электрических потенциалов (РПП и ПРП).
· ПРП тормозит выделение медиатора, что вызывает длительную гиперполяризацию мембраны биполяров и горизонтальных клеток.
· Суммируя синаптические влияния определенного числа фоторецепторов, биполяры оказывают возбуждающее влияние на мембрану ганглиозных клеток, что приводит к генерации распространяющихся нервных импульсов.
· На «выходе» сетчатки пространственное распределение возбужденных и невозбужденных фоторецепторов трансформируется в «мозаику» возбужденных рецептивных полей ганглиозных клеток, которые передают информацию о пространственных параметрах раздражителя в центральные отделы зрительной системы.
Сетчатка как система позволяет выделять такие
характеристики светового сигнала, как его интенсивность (яркость),
пространственные параметры (размер, конфигурация). Рецептивные поля,
построенные по принципу антагонистических отношений центра и периферии,
позволяют оценивать контрастность и контуры изображения, а также оптимальным
образом выделять полезный сигнал из шума [17, 412 c.].
1.4 Зрительный нерв
Аксоны ганглиозных клеток сетчатки, выходя из глазного яблока, образуют компактный пучок нервных волокон - зрительный нерв. Аксоны ганглиозных клеток, собранные в зрительном нерве, направляются к основанию передней части гипоталамуса, где оба нерва сходятся вместе, образуя хиазму (перекрест). Здесь происходит частичный обмен волокнами с разделением их на перекрещивающиеся и неперекрещивающиеся пучки. Дальше зрительные пути снова расходятся в виде правого и левого зрительных трактов [12, 411 c.].
Представьте себе, что вы смотрите на зрительную систему человека сверху. С этой удобной позиции вы могли бы увидеть, что все аксоны ганглиозных клеток с той половины сетчатки, которая ближе к носу, переходят в области хиазмы на противоположную сторону. В результате информация обо всем, что проецируется на внутреннюю (носовую) половину сетчатки левого глаза, переходит в правый зрительный тракт, а о том, что проецируется на носовую часть сетчатки правого глаза, - в левый зрительный тракт. Информация же от наружных (височных) половин обеих сетчаток идет по не перекрещенным путям. После хиазмы все стимулы, относящиеся к левой стороне внешнего мира, воспринимаются правой половиной зрительной системы, и наоборот.
Объединение аксонов зрительных нервов в
зрительный тракт носит не случайный характер. Волокна перекрещиваются таким
образом, что аксоны из соответственных участков обеих сетчаток встречаются и
вместе направляются к таламусу. Когда вы смотрите прямо перед собой, все предметы,
не находящиеся на средней вертикали, попадают на рецептивные поля клеток
носовой (внутренней) половины сетчатки одного глаза и височной (наружной)
половины сетчатки другого глаза. Таким образом, каждая точка внешнего
пространства проецируется на соответственные (корреспондирующие) точки обеих
сетчаток. Дальнейшие отображения всей совокупности таких точек в зрительной
системе называются ретинотопическими проекциями поля зрения. Ретинотопическая
организация характерна для всей структуры зрительной системы
[13, 386 c.].
1.5 Наружное
коленчатое тело
Аксоны зрительного тракта подходят к одному из четырех воспринимающих и интегрирующих центров второго порядка. Ядра латерального коленчатого тела и верхних бугорков четверохолмия - это структуры-мишени, наиболее важные для осуществления зрительной функции. Коленчатые тела образуют «коленоподобный» изгиб, и одно из них - латеральное (т.е. лежащее дальше от срединной плоскости мозга) - связано со зрением. Бугорки четверохолмия - это два парных возвышения на поверхности таламуса, из которых верхние имеют дело со зрением. Третья структура - супрахиазменные ядра гипоталамуса (они расположены над зрительным перекрестом) - используют информацию об интенсивности света для координации наших внутренних ритмов. И наконец, глазодвигательные ядра координируют движения глаз, когда мы смотрим на движущиеся предметы.
Латеральное коленчатое ядро. Аксоны ганглиозных клеток образуют синапсы с клетками латерального коленчатого тела таким образом, что там восстанавливается отображение соответствующей половины поля зрения. Эти клетки в свою очередь посылают аксоны к клеткам первичной зрительной коры - зоны в затылочной доле коры.
Верхние бугорки четверохолмия. Многие аксоны ганглиозных клеток ветвятся, прежде чем достичь латерального коленчатого ядра. В то время как одна ветвь соединяет сетчатку с этим ядром, другая идет к одному из нейронов вторичного уровня в верхнем бугорке четверохолмия. В результате такого ветвления создаются два параллельных пути от ганглиозных клеток сетчатки к двум различным центрам таламуса. При этом обе ветви сохраняют свою ретинотопическую специфику, т. е. приходят в пункты, в совокупности образующие упорядоченную проекцию сетчатки. Нейроны верхнего бугорка, получающие сигналы от сетчатки, посылают свои аксоны к крупному ядру в таламусе, называемому подушкой. Это ядро становится все крупнее в ряду млекопитающих по мере усложнения их мозга и достигает наибольшего развития у человека. Крупные размеры этого образования позволяют думать, что оно выполняет у человека какие-то особые функции, однако истинная его роль пока остается неясной. Наряду с первичными зрительными сигналами нейроны верхних бугорков получают информацию о звуках, исходящих от определенных источников, и о положении головы, а также переработанную зрительную информацию, возвращающуюся по петле обратной связи от нейронов первичной зрительной коры. На этом основании полагают, что бугорки служат первичными центрами интегрирования информации, используемой нами для пространственной ориентации в меняющемся мире.
1.6 Зрительная кора
Кора имеет слоистую структуру. Слои отличаются друг от друга строением и формой образующих их нейронов, а также характером связи между ними. По своей форме нейроны зрительной коры делятся на большие и малые, звездчатые, кустовидные, веретенообразные [20, 87 c.].
Известный нейропсихолог Лоренте де Но в 40-х гг. двадцатого столетия обнаружил, что зрительная кора делится на вертикальные элементарные единицы, представляющие собой цепь нейронов, расположенных во всех слоях коры [16, 57 c.].
Синаптические связи в зрительной коре весьма многообразны. Кроме обычного деления на аксосоматические и аксодендрические, концевые и коллатеральные, их можно подразделить на два типа: 1) синапсы с большой протяженностью и множественными синаптическими окончаниями и 2) синапсы с малой протяженностью и одиночными контактами [2, 497 c.].
Функциональное значение зрительной коры чрезвычайно велико. Это доказывается наличием многочисленных связей не только со специфическими и неспецифическими ядрами таламуса, ретикулярной формацией, темной ассоциативной областью и т.д.
На основании электрофизиологических и нейропсихологических данных можно утверждать, что на уровне зрительной коры осуществляется тонкий, дифференцированный анализ наиболее сложных признаков зрительного сигнала (выделение контуров, очертаний, формы объекта и т.д.). На уровне вторичной и третичной областей, по-видимому, происходит наиболее сложный интегративный процесс, подготавливающий организм к опознанию зрительных образов и формированию сенсорноперцептивной картины мира.