Материал: Половые клетки и оплодотворение
Моно- и полиспермия
Проникновение в яйцеклетку одного сперматозоида, получило название, физиологической моноспермии. Моноспермия присуща всем группам животных с наружным осеменением и многим животным с внутренним осеменением (тем, которые подобно, млекопитающим имеют яйцеклетки небольшого размера).
У других животных, например, у
некоторых членистоногих (насекомые), моллюсков (класс брюхоногих), хордовых
(акулообразные рыбы, хвостатые амфибии, рептилии и птицы) в яйцеклетку
проникает большое количество сперматозоидов. Такое явление получило название
физиологической полиспермии. Однако и в этом случае с ядром яйцеклетки
соединяется только ядро одного сперматозоида, тогда как остальные разрушаются
(рис.11).
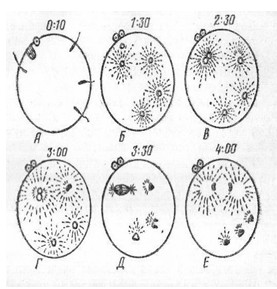
Рис. 11.
Полиспермия у тритона. А-проникновение спермиев в яйцо
на стадии метафазы II
деления созревания; Б-синхронные изменения семенных ядер, образование семенных
звезд; В-женское ядро соединяется с одним из семенных ядер; Г - Е-синкарион
вступает в митоз, сверхчисленные семенные ядра оттесняются в вегетативное
полушарие и дегенерируют. Цифры над изображением яиц - время после проникновения
спермиев при температуре 23о (по Гинзбург).
При физиологической моноспермии
имеются особые механизмы защиты яйца от полиспермии. Первый механизм связан с
изменением мембранного потенциала. Установлено, что в яйцеклетке лягушки, через
несколько секунд, после контакта со сперматозоидом заряд мембраны изменяется от
-28 до 8 мв и остается положительным в течение 20 мин. Такие же изменения
мембранного потенциала были обнаружены в яйцеклетках морского ежа. Оказалось,
что положительный заряд мембраны препятствует полиспермии. Другой широко
распространенный механизм защиты яйца от проникновения сверхчисленных
сперматозоидов связан с образованием оболочки оплодотворения и перивителлиновой
жидкости.
4. Партеногенез, гиногенез, андрогенез. Естественный и искусственный партеногенез
Яйцеклетки некоторых животных могут развиваться во взрослую особь и без оплодотворения. Такое развитие называется естественным партеногенезом или девственным развитием Оно встречается среди насекомых, низших ракообразных, коловраток, моллюсков и других животных. У многих из них партеногенез чередуется с половым размножением. Так, в течение лета коловратки откладывают яйца (летние яйца), из которых развиваются без оплодотворения только женские особи. Последнее поколение партеногенетических яиц дает и самцов и самок, откладывающих уже оплодотворенные яйца.
Пчелы, осы, муравьи, откладывают яйца, из которых часть оплодотворенные, а часть - неоплодотворенные. Из оплодотворенных яиц развиваются самки, а из неоплодотворенных - самцы. Такое своеобразное явление получило название факультативного партеногенеза.
Некоторые животные, обычно развивающиеся из неоплодотворенных яиц, начинают размножаться уже на личиночной стадии. Это яаление называют педогенезом. Оно встречается у некоторых мух, сосальщиков, ракообразных и других.
Партеногенетическое развитие широко распространено не только среди беспозвоночных, но и у позвоночных животных (рыбы, птицы, пресмыкающиеся). Так, у рыб партеногенетические яйца дают нормальных особей, но только самок. Из неоплодотворенных яиц развиваются нормальные особи мужского пола. Скальные ящерицы, содержащиеся в лабораторных условиях отдельно от самцов, в течение двух лет откладывали яйца, из которых развивались только самки. У других видов позвоночных естественный партеногенез неизвестен.
Наблюдаемые явления партеногенетического развития изучены с генетической и эмбриологической стороны еще недостаточно. Известно, что у пчел и ос, развивающихся партеногенетически, соматические клетки содержат гаплоидный набор хромосом (гаплоидный партеногенез). В большинстве случаев после партеногенетической активации яйцеклетки вней восстанавливается диплоидный набор хромосом. Существует два основных способа диплоидизации: амейотический и мейотический. При аймеотическом партеногенезе выпадает процесс редукции числа хромосом. При мейотическом партеногенезе редукция числа хромосом происходит, но диплоидный набор восстанавливается путем объединения хромосомных наборов обоих ядер, получившихся в результате митотического деления гаплоидного ядра яйцеклетки. Таким образом, в соматических клетках зародыша сохраняется диплоидный набор хромосом.
С открытием естественного партеногенеза были предприняты попытки искусственно вызвать развитие яиц. Первые опыты по искусственному партеногенезу были проведены А.А. Тихомировым в 1886 году на тутовом шелкопряде. Исследователем было обнаружено, что неоплодотворенные яйца тутового шелкопряда начинают развиваться после обработки их концентрированной серной кислотой, при изменении температуры или после механического повреждения, вызываемого потиранием яиц щеткой.
Обширные исследования были проведены Ж. Лебом в 1913, 1916 гг. Он показал, что если яйца морского ежа поместить в масляную, уксусную или в валериановую кислоту на несколько минут, а затем последовательно перенести в подщелоченную воду и нормальную воду, то наступала активация яйцеклетки.
Большой интерес вызвали опыты Е. Батайона в 1910 г. Исследователь вызвал партеногенетическое развитие лягушек и жаб уколом иглы.
Ж. Принкусу в 1956 г. удалось добиться партеногенетического развития яиц млекопитающих (кроликов). Известны многие попытки вызвать партеногенетическое развитие яйцеклеток других млекопитающих. Однако достоверных данных о возможности партеногенетического развития у млекопитающих на более поздних стадиях, чем бластоциста, пока нет.
Из изложенного выше видно. Что партеногенетическое развитие яиц можно вызвать воздействием на них разнообразными механическими, физическими и химическими агентами. Действия данных факторов приводит к одним общим изменениям, подобным тем, которые происходят в яйцеклетке при оплодотворении.
Опыты по искусственному партеногенезу не только способствуют выяснению механизма этого процесса, но и приобретают большое практическое значение. Б.Л. Астауров в 40-х годах разработал промышленный способ стимулирования развития неоплодотворенных яиц тутового шелкопряда при действии высоких температур. Этот способ позволяет получать в большом количестве партеногенетических бабочек женского пола и тем самым значительно увеличить производство коконов. Позднее З.А. Струнников разработал метод низкотемпературной активации яиц. Партеногенетическое потомство, полученное этим способом, состоит из одних самцов, которые вырабатывают больше шелка, чем самки. Проблема партеногенеза ждет новых исследователей, так как представляет большой теоретический и практический интерес.
Гиногенез и андрогенез
Гиногенез - явление сходное с партеногенезом и отличается от него тем, что при гиногенезе сперматозоиды участвуют как активаторы развития яйцеклетки, не внося свой генетический материал в яйцеклетку. Например, яйца серебряного карася могут быть стимулированы спермой сазана, плотвы, обыкновенного карася. Развитие зародыша осуществляется только за счет женского ядра. В популяциях гиногенетических животных встречаются только особи женского пола. Гиногенез обнаружен у круглых червей, серебряного карася и некоторых других животных. Гиногенетическое развитие можно вызвать искусственно, воздействуя на сперматозоиды рентгеновскими лучами, температурой и другими агентами.
Андрогенез - явление противоположное гиногенезу. При андрогенезе развитие осуществляется за счет ядер сперматозоидов и цитоплазмы яйцеклетки. Андрогенез наблюдается в тех случаях, когда ядро яйцеклетки погибает, а в яйцеклетку проникает несколько спермиев. Ядра двух сперматозоидов объединяются, и образуется ядро с диплоидным набором хромосом. Естественный андрогенез обнаружен у тутового шелкопряда и паразитической осы. При андрогенезе образуются только самцы. Андрогенез может быть вызван и искусственно. Особую известность приобрели исследования Б.Л. Астаурова (1957) по искусственному андрогенезу у тутового шелкопряда, имеющие как теоретическое, так и практическое значение. Эти способом можно получить популяцию самцов тутового шелкопряда, коконы которых содержат на 25-30% шелка больше, чем коконы самок. Для получения андрогенетических особей, яйца тутового шелкопряда облучались большой дозой рентгеновских лучей, при этих условиях ядро разрушалось, а ооплазма не утрачивала жизнеспособности. После этого осуществляли искусственное осеменение. Два из многих, проникших в клетку сперматозоида сливались и давали начало ядру с диплоидным набором хромосом. Такое ядро и обеспечивало нормальное развитие.
Пол зародыша обычно обусловлен генотипически и зависит от сочетания половых хромосом в оплодотворенном яйце. При помощи половых хромосом регулируется численное равенство в природе.
Для обозначения половых хромосом у большинства млекопитающих, некоторых амфибий(леопардовая лягушка) и насекомых (дрозофила) используют латинские буквы Х и У. Половые хромосомы самки обозначают ХХ (знак ♀), а половые хромосомы самца ХУ (знак ♂). В этом случае самцы продуцируют два типа сперматозоидов Х - или У - хромосомой. У самок же образуется один тип яиц с Х-хромосомой. Особи, образующие один тип половых клеток называются гомогаметными, а те особи, которые образуют два типа половых клеток - гетерогаметными, т.е. самки были гомогаметны, а самце - гетерогаметны.
Если яйцо будет оплодотворено сперматозоидом с Х-хромосомой, из него разовьется самка, содержащая ХХ-хромосомы. Если же яйцо будет оплодотворено сперматозоидом с У-хромосомой, то из него образуется самец, содержащий ХУ-хромосомы. Таким образом, у перечисленных выше животных У-хромосома содержит гены, которые определяют мужской пол.
У некоторых животных (бабочки, рыбы, птицы) гетерогеметен женский пол. В этом случае для обозначения половых хромосом используют другие буква - Z и W. Половые хромосомы самки обозначают буквами ZW, а половые хромосомы самца - ZZ. У этих животных самки продуцируют два типа яйцеклеток с Z или W-хромосомой., а самцы - один тип сперматозоидов с Z - хромосомой.
В этих механизмах нет ничего принципиально различного, за исключением того, что гетерогаметны в одном случае самцы, а в другом самки.
Полученные в последние годы экспериментальные данные показали, что гены, ответственные за определение пола, располагаются не только в половых хромосомах, но и в аутосомах. Тл обычно является результатом взаимодействия нескольких пар генов. Пол является полигенным признаком.
Следует также отметить, что
рассмотренный выше хромосомный механизм определения пола фиксирует лишь
первичные признаки пола - строение половых жлез, в то время как вторичные
половые признаки при определенных условиях могут претерпевать обратное
развитие, превращаясь в признаки противоположного пола. Решающую роль в этом
играют половые гормоны.
Литература
яйцеклетка сперматозоид партеногенез гиногенез
Биология. Пособие для поступающих в вузы / А.Г. Мустафин, Ф.К. Лагнуев, Н.Г. Быстренина и др., под ред. В.Н. Ярыгина. - М.: Высшая школа, 2005. - 492 с.
Биология. Справочник студента / А.А. Каменский, А.И. Ким, Л.Л. Великанов, О.Д. Лопина, С.А. Баландин, М.А. Валовая, Г.А. Беляков. - М.: Физиологическое общество «СЛОВО», 2007. - 640 с.
Биология. Справочник школьника и студента / Под ред. З. Брема, И. Мейнке. - М.: Дрофа, 2004. - 400 с.
Вахненко Д.В., Гарнизоненко Т.С., Колесников С.И. Биология с основами экологии. Учебник для вузов / Д.В. Вахненко, Т.С. Гарнизоненко, С.И. Колесников. - Ростов-на-Дону: Феникс, 2008. - 448 с.
Грин Н., Стаут У., Тейлор Д. Биология / Н. Грин, У. Стаут, Д. Тейлор, под ред. Р. Сопера. - М.: Мир, 2004. - Т. 1. - 368 с.
Грин Н., Стаут У., Тейлор Д. Биология / Н. Грин, У. Стаут, Д. Тейлор, под ред. Р. Сопера. - М.: Мир, 2004. - Т. 2. - 325 с.
Грин Н., Стаут У., Тейлор Д. Биология / Н. Грин, У. Стаут, Д. Тейлор, под ред. Р. Сопера. - М.: Мир, 2004. - Т. 3. - 325 с.
Кемп П., Армс К. Введение в биологию / П. Кемп, К. Армс. - М.: Мир, 2000. - 671 с.
Лысов П.К., Акифьев А.П., Добротина Н.А. Биология с основами экологии: Учебник/ П.К.Лысов, А.П.Акифьев, Н.А.Добротина- М.: Высшая школа., 2007.- 655 с.
Пехов А.П. Биология с основами экологии. Учебное пособие для вузов с грифом МО / А.П. Пехов. - СПб.: Изд-во «Лань», 2008. - 672 с