Материал: Основные понятия и определения в иммунологии, Годовалов, Южанинова, 2017

6. Рецепторы для распознавания антигенов
6.1. Теория "боковых цепей"
Пауль Эрлих ‒ автор теории "боковых цепей":
1.В основе реакции антиген-антитело лежат законы структурной химии
2.Антитела уже предсуществуют в организме до его контакта с антигеном и находятся на поверхности клеток в виде "боковых цепей" или рецепторов
Пауль Эрлих
1854–1915
Основные положения:
1. В основе реакции антиген-антитело лежат законы структурной химии или комплементарные взаимодействия, что определяет специфичность иммунных реакций.
По сути, это другое приложение принципа "ключа и замка", сформулированного выдающимся биохимиком Эмилем Фишером для объяснения механизма работы ферментов. По представлениям Э. Фишера в основе специфичности любой ферментативной реакции лежит полное пространственное соответствие в расположении функциональных групп активного центра фермента и субстрата. Иначе говоря, субстрат подходит к активному центру фермента, как ключ подходит к замку. П. Эрлих использовал эту идею для объяснения механизма реакции антиген-антитело: антигенные детерминанты подходят к активному центру антител как ключ к замку, т.е. в основе взаимодействия антигена и антитела лежит пространственная комплементарность молекул. Стоит добавить, что взаимодействие субстрат/фермент и антиген/антитело ‒ это частные проявления универсального биологического принципа специфичности взаимодействия любого рецептора с его лигандом.
35
Согласно современным представлениям, для того, чтобы антитело реализовало свои функции, необходима точная пространственная стыковка антигенраспознающего центра и эпитопа. Такая стыковка возможна в том числе благодаря подвижности шарнирного участка антител, соединяющего Fab-фрагменты. Точное взаимодействие антиген-антитело запускает конформационные изменения в Fcфрагменте антител, а это, в свою очередь, приводит к запуску эффекторных функций антител.
2. Антитела уже предсуществуют в организме до его контакта с антигеном и находятся на поверхности клеток в виде "боковых цепей" или рецепторов. Антиген, попавший в организм, в соответствии с законами структурной химии связывается лишь с тем рецептором, который комплементарен ему. Это приводит к избирательной активации синтеза лишь данного типа антител.
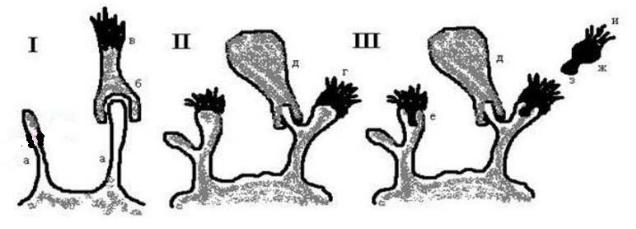
Оригинальный рисунок П. Эрлиха, иллюстрирующий положения его теории
I – рецептор 1-го типа: а – чувствительный комплекс, б – чувствительная группа, в - токсин II – рецептор 2-го типа: г – активный центр ферментной группы, д – адсорбированная мккула
III – рецептор 3-го типа: е – комплект, воспринимающий комплементарные молекулы, ж – комплементарная молекула, з – чувствительная группа, и – ферментотоксическая группа.
36
6.2. Клонально-селекционная теория (Ф.М. Бернет, конец 50-х годов XX века)
Основные постулаты:
1.Клональная гетерогенность или клональное разнообразие лимфоцитов
Ворганизме человека присутствует 1012 лимфоцитов, которые сгруппированы в клоны. Клон - популяция клеток, происходящая из одного предшественника путем деления. Все потомки клетки-предшественника генетически идентичны. Следовательно, все клетки одного клона имеют одну специфичность, т.е. одинаковую структуру своих антигенраспознающих рецепторов.
Формирование клональной гетерогенности происходит до момента встречи с антигеном на этапе антигеннезависимой дифференцировки лимофитов. В основе формирования репертуара антигенраспознающих рецепторов лежит процесс реаранжировки генных сегментов (случайное соединение V-, D-, J-сегментов генов антигенраспознающих рецепторов) и соматические мутации в тех же участках гена.
2.Поступление в организм малых количеств антигена приводит к связыванию его с рецепторами соответствующих клонов лимфоцитов. Это приводит к активации соответствующих клонов с последующим вступлением их в процессы пролиферации (в результате чего численность клона увеличивается в геометрической прогрессии) и дифференцировки в соответствующие эффекторные клетки и клетки-памяти.
3.Введение большого количества антигена приводит к формированию иммунологической толерантности, т.е. специфической иммунной ареактивности, связанной с элиминацией (гибелью) клонов, несущих рецепторы к данному антигену. Это лежит в основе формирования естественной врожденной толерантности к своим собственным антигенам.
Ссовременных позиций в основе формирования толерантности могут лежать и другие механизмы, например, специфическая супрессия отвечающих клеток регуляторными Т-лимфоцитами. Механизм направленной гибели клонов играет важную роль в отрицательной селекции клонов на этапе антигеннезависимой дифференцировки.
37
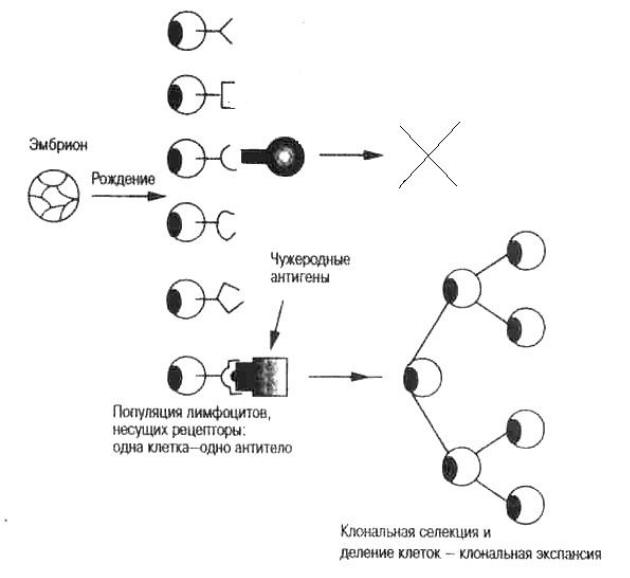
Схематическое изображение теории Ф.М. Бернета
38
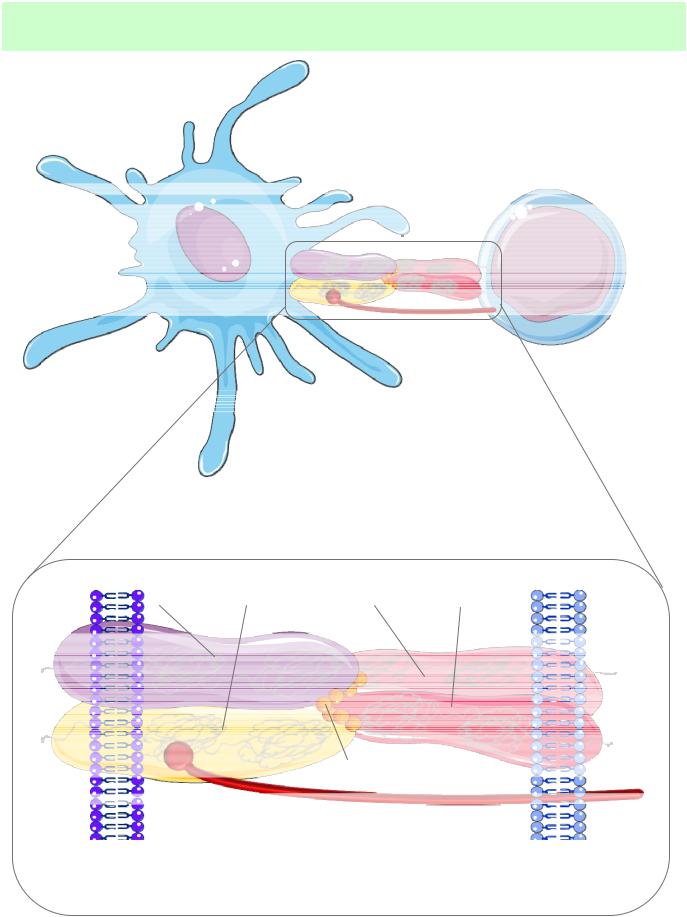
7. Примеры взаимодействия компонентов палео- и неоиммунитета
Распознавание антигена в виде комплекса пептид-МНС II ТCR
дендритная клетка
CD4+ Т-лимфоцит
молекула MHCII: |
Т-клеточный рецептор: |
||
α-цепь и β-цепь |
α-цепь и β-цепь |
||
|
|
|
|
|
пептид |
|
молекула CD4 |
мембрана денд- |
мембрана |
ритной клетки |
CD4+ T-клетки |
39