Материал: Общая характеристика бактерий рода BACILLUS
аспорогенная формы культур одного и того же вида различны. Не установлено существование какого-либо антигена, общего для спор бактерий разных видов.
Наоборот, по антигенной структуре имеется достаточно отчетливая дифференциация между спорами разных видов.
В способе прорастания споры и продолжительности процесса также отмечается определенное постоянство, характерное для различных видов бактерий.
Старые споры прорастают в течение более длительного периода, чем молодые. Время прорастания молодых спор является довольно постоянным у различных бактерий и может служить видовым признаком. Если перенести только что сформировавшиеся споры в благоприятную питательную среду, то прорастание их начинается не сразу, а по истечении 2-3 ч.
Установлено, что на начальной стадии прорастания спор происходит выделение в среду так называемого экссудата прорастания,
характеризующегося специфическим поглощением ультрафиолетовых лучей.
Вещество это было выделено и идентифицировано как кальциевая соль дипиколиновой кислоты (пиридин - 2,6-дикарбоксиловая кислота).
Дипиколинат кальция составляет приблизительно половину сухой массы экссудата прорастания, что соответствует 15% от общей сухой массы покоящихся спор. Остальная часть экссудата прорастания представлена свободными аминокислотами, высокомолекулярными и низкомолекулярными белковыми веществами. Обнаружено особенно большое количество не диализируемого гексозаминсодержащего пептида с молекулярным весом порядка 10 000, в состав которого входят диаминопимелиновая кислота,
аланин, глютаминовая кислота и гексозамины. Условно это соединение названо ДАП-гексозаминпептид. Это соединение входит в состав оболочки спор, в
отличие от дипиколиновой кислоты, обнаруживаемой внутри спор.
Среди факторов, способствующих процессу прорастания спор, большое внимание уделяется тепловой активации, действие которой более выражено при наличии в среде некоторых химических соединений. Тепловая активация
16
достигается предварительным кипячением суспензии спор при 97-100° C в
течение 10-15 мин или при 65° C в течение 2 ч. Иногда этот эффект достигается при обычной пастеризации. Тепловая активация прорастания спор носит общий характер для разных видов спороносных бактерий, хотя она более специфична для аэробов, чем анаэробных видов. Обычно тепловое воздействие усиливает и ускоряет прорастание спор в течение короткого промежутка времени. Действие подобной обработки спор, как правило, преходяще, хотя при этом отмечается и некоторое изменение условий, необходимых для течения процесса прорастания спор. Так, для лучшего прорастания спор сенного бацилла без предварительной тепловой обработки необходимо присутствие в среде L-аланина, аденозина,
глюкозы и тирозина, тогда как после тепловой обработки оптимальные условия прорастания спор обеспечивают наличие лишь глюкозы и аденозина.
Установлено также, что сохранение в течение длительного времени спор при
37° C после их тепловой активации ведет к деактивации. Тепловая обработка этих спор после длительного хранения вновь ведет к реактивации прорастания.
Имеются данные о некотором активирующем влиянии на прорастание спор умеренного воздействия ультразвука.
Исследования последних лет показали, что интенсивность прорастания спор специфически повышается в присутствии некоторых веществ, в первую очередь L-аланина, пуриновых нуклеозидов и глюкозы. Иногда необходимо наличие, помимо L-аланина, ряда других аминокислот. Из рибозидов наиболее эффективным оказался аденозин.
Аланин является наиболее специфическим активатором процесса прорастания спор из всех известных соединений, обладающих подобным действием.
Оптимальные условия для прорастания спор анаэробных бактерий несколько более сложны, хотя факторы, активирующие процессы прорастания спор аэробных видов, и в этом случае оказывают активирующее действие.
Наряду с активаторами выявлен ряд веществ, угнетающих процесс прорастания спор. Прорастание спор у аэробных видов угнетается в
17
присутствии оксина, некоторых мышьяковистых соединений, углекислого натрия, высоких концентраций ряда аминокислот - глицина, метионина,
цистеина и валина. Характерно угнетающее действие D - аланина в отличие от
L-аланина.
Механизм активирующего и угнетающего действия ряда факторов на прорастание спор остается неясным. Одни авторы приписывают активаторам определенную роль в процессах деполимеризации различных составных элементов в спорах, высвобождении и активации ряда ферментных систем,
обусловливающих начало прорастания спор. Другие считают их энергетически активными соединениями, стимулирующими биохимические процессы в момент прорастания спор.
Обычно не все споры, вносимые в благоприятную среду, прорастают сразу. Некоторое их количество остается не проросшими в течение различного времени, не теряя, однако, своей жизнеспособности. Установлено, что при длительном хранении процент прорастающих спор постепенно снижается.
Причина этого явления и природа подобных «дремлющих» спор остаются неясными.
В покоящейся споре имеются ферментные системы, выявляемые лишь после их активации под влиянием определенных факторов и химических соединений, которые, однако, исключают прорастание спор. К таким системам относятся прежде всего окислительные ферменты. Активация их достигается обработкой спор незначительными количествами L-аланина, аденозина и некоторыми другими веществами, а также кипячением или пастеризацией суспензий спор. Перевод неактивной ферментной системы в активную без признаков прорастания спор считается первым этапом биохимических изменений в спорах, ведущих в дальнейшем к началу процесса их прорастания.
Полагают, что под действием активаторов дипиколиновая кислота,
связывающая окислительные и другие ферменты, подвергается деполимеризации с освобождением ферментных систем.
18
С помощью высокочувствительного микрореспирометра у бактериальных спор установлена дыхательная активность.
При изучении химического состава клеточной стенки спор различных видов аэробных бактерий обнаружены характерные различия в содержании ДАП-
гексозаминпептида и других веществ - полисахаридов, липидов. ДАП-
гексозаминпептид высвобождается из стенки спор под действием литического агента, уподобляемого по своим свойствам лизоциму (Жизнь растений.
Энциклопедия в шести томах. Том 1. Введение. Бактерии и актиномицеты / Под ред. чл. - кор. АН СССР, проф. Н. А. Красильникова и проф. А. А. Уранова, 1974).
Стадии споруляции. Бактерии достаточно сложно принять решение о начале споруляции, поскольку этот процесс, будучи запущенным, уже необратим. А поскольку это решение коренным образом меняет метаболизм клетки, ошибка может иметь для нее печальные последствия. Поэтому для принятия этого решения бактерия должна проанализировать множество сигналов от внешней среды от внутриклеточных метаболических систем, от клеточного цикла. Сигналы могут происходить от нехватки питательных веществ, плотности популяции, синтеза ДНК, повреждений ДНК, цикла Кребса
(Hoch,1978).
Споруляция разделяется на семь стадий, различия между которыми основаны на цитологических изменениях, происходящих в процессе образования споры. Стадия 0, являющаяся подготовительной стадией споруляции, наблюдается в конце логарифмической фазы роста, или в период То (Т1, Т2, Т3 и т. д. соответствуют времени, в часах, по отношению к концу логарифмической фазы). В лабораторных условиях процесс споруляции обычно занимает около 6-8 ч, и, таким образом, порядковый номер стадии развития и время, выраженное в часах, почти совпадают: например, стадия IV наблюдается в периоде Т4.
19
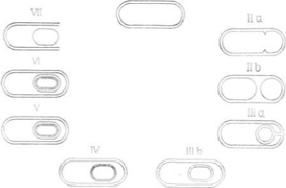
Рис 2.Стадии споруляции у В. sublilis. Последовательность событий приведена в направлении по часовой стрелке от начальной стадии 0 в конце логарифмической фазы роста.
На стадии 0 никаких морфологически различимых споруляционных структур нет, и клетки имеют вид, характерный для вегетативной стадии. В
настоящее время на стадии I не идентифицированы какие-либо морфологические изменения, характерные для процесса споруляции; таким образом, можно считать, что развивающиеся клетки переходят от стадии 0 к
стадии II.
Стадия II - это первая стадия споруляции, распознаваемая морфологически. Она характеризуется появлением проспоровой перегородки,
которая асимметрично делит клетку на маленькую проспору и большую материнскую клетку. Это критический шаг, так как образование асимметрично расположенной мембраны является ключевым этапом споруляции.
В течение стадии III материнская клетка поглощает проспору. Проспора,
которая теперь находится в цитоплазме материнской клетки, имеет на периферии двойную мембранную структуру, и молекулы, перемещающиеся из цитоплазмы материнской клетки в цитоплазму проспоры, должны пройти через эту двойную мембрану по следующему пути:
Цитоплазма материнской клетки→ Внешняя мембрана проспоры (изнутри-
наружу)→пространство между мембранами Внутренняя мембрана проспоры (снаружи внутрь) → цитоплазма проспоры
20