Материал: Общая характеристика бактерий рода BACILLUS
состоят из чередующихся L- и D-аминокислот, причем наиболее часто встречается последовательность L-аланин/D-глутаминовая кислота/L-
диаминокислота (обычно м-диаминопимелиновая кислота или L-лизин). Если у грамотрицательных бактерий химический состав и структура пептидогликана достаточно однообразны, и, следовательно, имеют небольшую ценность для целей классификации, то у грамположительных бактерий обнаруживается большое разнообразие состава и структуры пептидных цепей, которые могут соединяться поперечными сшивками непосредственно или через межпептидные мостики. Было описано около 100 различных типов пептидогликанов. Поскольку структура и состав пептидогликанов, образуемых различными бактериями, обычно стабильны и не зависят от условий культивирования, то состав клеточной стенки у многих грамположительных бактерий часто служит важным хемотаксономическим признаком.
У большинства видов Bacillus муреин представлен вариантом с пептидами, непосредственно соединенными через мезодиаминопимелиновую кислоту (mDpm или mА2pm) (Харвуд, 1992).
1.3 Строение спор
Поверхность наружной оболочки спор разнообразна у аэробных бактерий: либо гладкая, либо с выростами, выступами и почками. Оболочка составляет значительную часть споры. Основными ее компонентами являются белки (60-90%) и липиды. Состав аминокислот в белках оболочки неодинаков у бактериальных спор разных видов, однако большая часть цистина,
содержащегося в значительном количестве в спорах, обнаруживается именно в оболочке.
11
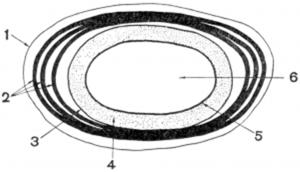
Рис 1. Схемотическое строение споры:
1 -экзоспориум; 2 -слои споровой оболочки; 3 - внешняя мембрана споры; 4 - кора; 5 - внутренняя мембрана споры; 6 — сердцевина.
Под оболочкой располагается кора споры, состоящая в основном из муреина, который является нерастворимым структурным материалом коры.
В течение многих лет покоящаяся спора рассматривалась как почти инертное тело, обладающее несколькими ферментами или вовсе лишенное их.
Некоторых ферментных систем, имеющихся в вегетативной клетке, например цитохромов и ферментов цикла трикарбоновых кислот, ничтожно мало в споре.
Это указывает на то, что в спорах аэробный энергетический метаболизм осуществляется иным путем, чем в клетках. Аналогичные ферменты спор и вегетативных клеток в спорах более термоустойчивы.
Споры аэробных бактерий обладают различной термоустойчивостью, что обусловлено как видовыми особенностями, так и условиями их образования.
Установлено, что споры, образующиеся при высокой температуре, а также полученные на средах с высоким содержанием кальция, парааминобензойной кислоты и некоторых жирных кислот, более жароустойчивы.
Споры более устойчивы, чем вегетативные клетки, от которых они образовались, к воздействию проникающей радиации, ультразвука,
гидростатического давления, замораживания, разрежения и др. Устойчивость спор к кислотам и щелочам во многом зависит от их принадлежности к различным видам бактерий. Споры некоторых культур бактерий остаются жизнеспособными даже при кипячении в концентрированной соляной кислоте в течение 20 мин. При этом резистентность спор во много раз повышается при
12
их обезвоживании (Жизнь растений. Энциклопедия в шести томах. Том 1.
Введение. Бактерии и актиномицеты / Под ред. чл. - кор. АН СССР, проф. Н. А.
Красильникова и проф. А. А. Уранова, 1974).
К спиртам и другим органическим растворителям устойчивость спор намного выше, чем у вегетативных клеток. Споры менее устойчивы к действию алкилирующих агентов. При воздействии комплекса определенных ферментов удается достигнуть растворения спор.
Механизм устойчивости спор к действию высокой температуры,
проникающей радиации и других физико-химических факторов полностью еще не выяснен. Термоустойчивость спор объясняется наличием кальция,
дипиколиновой кислоты и в некоторой степени диаминопимелиновой кислоты.
Эти вещества определяют в значительной мере и резистентность спор к действию радиации. Полагают, что споры содержат комплекс особых веществ,
подобных хелатным соединениям, которые предохраняют жизненно важные структурные элементы от губительного воздействия физико-химических факторов.
Важным фактором, приводящим к образованию спор, является отсутствие для данного вида аэробных бактерий благоприятных условий для роста и развития. Способствует спорообразованию обеднение питательной среды.
У аэробных бактерий анаэробные условия выращивания подавляют как спорообразование, так и рост. Спорообразование аэробных бактерий протекает лишь в условиях доступа кислорода. Интенсивность споруляции подавляется в разной степени при выращивании бактерий в неблагоприятных условиях - при температуре выше оптимальной или реакции среды менее подходящей для обильного развития. Неблагоприятные условия роста, ускоряющие споруляцию бактерий, как правило, не обусловливают обильного «урожая» спор (Жизнь растений. Энциклопедия в шести томах. Том 1. Введение. Бактерии и актиномицеты / Под ред. чл. - кор. АН СССР, проф. Н. А. Красильникова и проф. А. А. Уранова, 1974).
13
Разные виды аэробных бактерий отличаются расположением и формой спор. Эти признаки имеют важное систематическое значение. У некоторых видов расположение спор строго определенное - центральное, околоконцевое и концевое. У других видов не наблюдается строгой локализации.
Высокое содержание в питательной среде источников азотного и углеродного питания, обеспечивающих максимальный рост бактерий, может подавлять индукцию спорообразования. Так, потребность в глюкозе - основном энергетическом источнике вегетативного роста аэробных бактерий - является несущественной для спорообразования некоторых видов, а в отдельных случаях она угнетает процесс споруляции. Культуры определенных видов бактерий более интенсивно спорулируют при уменьшении в среде аланина, валина,
лейцина и изолейцина. У других бактерий отсутствие в среде лейцина и серосодержащих аминокислот угнетает спорообразование.
Интенсивность процесса образования спор в значительной мере обусловлена наличием в среде минеральных солей, содержащих ионы кальция,
магния, калия, марганца, железа, цинка, меди и некоторых редких металлов
(кобальта, лития, кадмия, никеля). Оптимальная концентрация минеральных солей и металлов различна в зависимости от вида выращиваемой культуры бактерий, условий ее роста. Роль минеральных солей и отдельных ионов металлов настолько велика, что в их отсутствии спороношение может вовсе не наблюдаться. Изменения концентраций их в питательной среде могут оказать иногда резко различное действие.
Спорообразование отдельных культур бактерий настолько зависит от наличия минимальных концентраций некоторых металлов, что эта особенность используется в аналитических целях.
Роль минеральных солей различных металлов в образовании бактериальных спор установлена недавно, и многие вопросы механизма их действия пока не выяснены. Полагают, что их специфическое действие связано с активированием различных ферментативных систем. Значение таких элементов, как кальций, заключается также и в том, что они способствуют
14
образованию разнообразных низкомолекулярных веществ в спорах,
обеспечивающих их термостабильность.
Сравнительно слабо изучено влияние на образование спор различных витаминов и других ростовых веществ. Некоторое усиление этих процессов отмечено под влиянием парааминобензойной и фолиевой кислот.
В результате всестороннего изучения биохимии спорообразования показано, что при споруляции не образуются новые ферментные системы.
Аналогичные ферменты, обнаруживаемые в вегетативной клетке и споре,
имеют одинаковые молекулярный вес, аминокислотный состав и другие физико-химические свойства. Теплоустойчивость ферментов спор вызвана присутствием в них ионов кальция, марганца и других металлов (Жизнь растений. Энциклопедия в шести томах. Том 1. Введение. Бактерии и актиномицеты / Под ред. чл. - кор. АН СССР, проф. Н. А. Красильникова и проф. А. А. Уранова, 1974).
Данные исследований свидетельствуют о том, что природа и функция ферментативного аппарата, осуществляющего синтез белков в споре и материнской вегетативной клетке, одинаковы. Основной системой транспорта электронов в спорах является растворимая флавопротеиноксидаза. При споруляции выявлена высокая активность ряда ферментов, например аргиназы,
некоторых нуклеаз. Основная масса белков споры синтезируется заново в период споруляции. Одновременно отмечается процесс распада белков, причем многими авторами обнаружена тесная взаимосвязь протеолитической активности с функцией спорообразования у бактерий. Из вновь синтезированных белков лишь незначительная часть используется для создания структурных элементов споры. В целом споруляцию можно охарактеризовать как процесс интенсивного распада белков и всех основных типов РНК материнской клетки с одновременным образованием новых белковых компонентов.
Различия в составе белков споры и вегетативной клетки выявляются серологическими методами. По антигенной структуре спорулирующая и
15