Материал: Молекулярно-генетическая характеристика рекомбинантных форм вируса иммунодефицита человека 1 типа, выявленных на территории Республики Беларусь
На 5’- и 3’-концах вирусной РНК находятся длинные концевые повторы (long terminal repeat, LTR), обеспечивающие инициацию, регуляцию и терминацию транскрипции, регуляцию упаковки генома в капсид и некоторые другие функции.
.3 Структура вириона ВИЧ
Вирусные частицы ВИЧимеют сферическую форму размером 100-120 нм, окруженную липидной мембраной(рисунок 1.2).
Липидная мембрана образована из клеточной мембраны клетки-хозяина, в
которой происходила репликация вириона. Каждая вирусная частица содержит 72
гликопротеиновых комплекса gp160, интегрированных в липидную мембрану [16].
Данные комплексы являются рецепторами вируса и состоят из двух вирусных белков:
поверхностного gp120 и трансмембранного gp41. К внутренней поверхности мембраны
прикреплен матриксный белок p17.
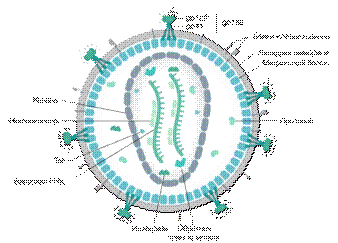
Капсид ВИЧсостоит из белка p24 и имеет коническую форму, суженная часть которого взаимодействует с оболочкой вириона. Внутри капсида находятся две молекулы однонитевой РНК (ssRNA), которые стабилизированы нуклеокапсидным белком p7. В капсиде также инкапсулированы вирусные ферменты: обратная транскриптаза, интеграза и протеаза, некоторые вспомогательные белки ВИЧ.
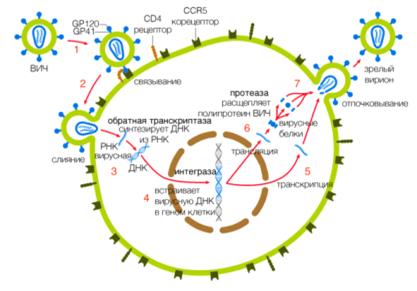
.4 Жизненный цикл ВИЧ
Жизненный цикл ВИЧможно разделить на несколько этапов(рисунок 1.3):
Связывание с рецепторами и корецепторами клетки-мишени
В начале исследований ВИЧ-инфекции был выявлен главный рецептор, с которым связывается вирус, - это гликопротеин CD4 (cluster of differentiation 4), найденный на поверхности иммунных клеток (Т-хелперов, макрофагов, моноцитов и дендритных клеток). Внедрение в клетку-мишень обеспечивается связыванием трехмерного гликопротеина ВИЧgp120 одновременно с рецептором CD4 и корецептором CCR5 или CXCR4 [17, 18].
Гликопротеин gp120 ВИЧсостоит из ~500 аминокислот и включает в себя 5 константных (C1-C5) и 5 вариабельных (V1-V5) областей различной длины. Домен связывания с CD4 находится в нескольких эпитопах областей V1-V2 и С2, C3, C4 и C5 [19].
Вначале происходит связывание CD4-связывающего домена gp120 с рецептором CD4. Это приводит к структурным изменениям молекулы gp120, открывая хемокин-связывающий домен. Данный домен, в свою очередь, связывается с одним из корецепторов клетки-мишени, что является сигналом для начала конформационных изменений трансмембранного белка gp41.
Проникновение ВИЧв клетку-мишень
После связывания gp120 ВИЧс рецептором CD4 и одним из корецепторов конформационные изменения в gp41 приводят к внедрению N-терминального гидрофобного региона этого протеина в мембрану клетки-мишени [20]. Результатом данного внедрения является слияние мембран вируса и клетки-мишени и, как следствие, попадание компонентов ВИЧв цитоплазму клетки [21].
После проникновения компонентов вируса в цитоплазму клетки вирусная РНК и ферменты находятся внутри матрикса, который разрушается под действием p38 MAP-киназы (MAPK) [22] и нуклеокапсида, разрушаемого циклофилином А (CypA) [23]. В результате этого генетический материал и ферменты вируса попадают в цитоплазму клетки-мишени, что приводит к инициации обратной транскрипции.
Обратная транскрипция
В цитоплазме клетки вирусная обратная транскриптаза (ОТ) при помощи фермента РНКазы H преобразует вирусную РНК в двухцепочечную ДНК методом обратной транскрипции. Было высказано предположение, что обратная транскрипция может происходить и в вирионе до внедрения в клетку, однако данный процесс сильно ограничен низкой концентрацией дНТФ внутри вириона [24].
Переход вирусного генома из РНК в ДНК происходит с помощью механизма, устраняющего два препятствия, с которыми сталкивается вирус в процессе репликации. Во-первых, требование системы РНК-праймера исключает точное транскрибирование генома в ДНК от начала до конца, так как не было бы никакого способа, чтобы скопировать область связывания праймера. Во-вторых, поскольку новые вирусные геномы должны быть получены с помощью клеточной РНК-полимеразы II, сигналы, соответствующие прямому синтезу РНК, должны находиться до начала сайта инициации транскрипции за пределами области, подлежащей копированию. С этой целью ВИЧ, как и остальные ретровирусы, синтезируют в обоих 5’- и 3’-концах ДНК дополнительную копию последовательностей, присутствующих только один раз в РНК-геноме, U5 и U3. Таким образом,длинные концевые повторы (LTR) (U3-R-U5) содержат практически все cis-активные элементы, необходимые для событий, которые происходят на уровне ДНК, то есть интеграции и экспрессии провирусной ДНК[25].
Инициация обратной транскрипцииВИЧпроисходит путем связывания клеточной транспортной РНК tRNALys3, выступающей в качестве праймера [26], с сайтом связывания праймера (primer binding site, PBS), расположенным на расстоянии приблизительно 180 нуклеотидов от 5’-конца вирусной РНК(рисунок 1.4, A). После связывания происходит синтез участков R и U5 кДНК, образуя таким образом гетеродуплекс РНК/ДНК, который является субстратом для РНКазы H(рисунок 1.4, B). РНКаза H расщепляет 5’-конец вирусной РНК, высвобождая вновь синтезированную (-)цепь ДНК. Концы вирусной РНК являются повторяющимися последовательностями, называемыми R. Эти повторы действуют как мост, который позволяет вновь синтезированной (-)цепи ДНК переходить от 5’- к 3’-концу вирусной РНК(рисунок 1.4, С). Было показано, что вновь синтезированная (-)цепь ДНК может связываться с 3’-концом любой из двух молекул РНК, упакованных в геном ретровируса [27, 28].
После связывания (-)цепи ДНК с R-областью 3’-конца РНК происходит синтез полной (-)цепи ДНК(рисунок 1.4, D). РНКаза H расщепляет РНК ВИЧ, за исключением участка, богатого пуринами (полипуриновый тракт, polypurine tract, ppt)(рисунок 1.4, E). Данный участок является резистентным к РНКаза H и служит праймером для инициации синтеза (+)цепи ДНК на матрице (-)цепи ДНК. ОТ ВИЧсинтезирует не только копию (-)цепи ДНК, но также первые 18 нуклеотидов праймера tRNALys3. Как только tRNA была скопирована в ДНК (+)цепи, она становится субстратом для РНКазы Н. Большинство ретровирусов удаляют всю tRNA, но ОТ ВИЧявляется исключением. Она оставляет один рибонуклеотид аденин на 5’-конце (-)цепи ДНК [29]. Расщепление tRNALys3 высвобождает участок PBS (primerbindingsite, сайт посадки праймера) на (+)цепи ДНК, который комплементаренPBS (-)цепи ДНК. Эти два комплементарных участка связываются между собой (рисунок 1.4, F), и происходит достраивание (+)цепи ДНК(рисунок 1.4, G).
В процессе обратной транскрипции синтезируется ДНК, которая длиннее РНК-матрицы: оба конца ДНК содержат последовательности с каждого конца РНК (U3 от 3’-конца и U5 от 5'-конца). Таким образом, каждый конец вирусной ДНК имеет ту же последовательность, U3-R-U5, которая после интеграции в геном клетки-хозяина будет являться концами провируса ВИЧ.
Интеграция вирусного генома в геном клетки-хозяина
Синтезированная двухцепочечная провирусная ДНК формирует пре-интеграционный комплекс с белками клетки-хозяина, вирусными белками интегразой, vpr и матриксным белком p17. Сигнал ядерной локализации внутри p17 направляет комплекс в ядро клетки [30, 31]. В ядре клетки геном ВИЧв виде двойной спирали ДНК интегрируется в геном клетки-хозяина путем сплайсинга с помощью фермента интегразы ВИЧ[32]. Результаты исследований показали, что провирусная ДНК предпочтительно интегрируется в области кодирования активных генов клетки [33]. Интегрированная форма генома ВИЧизвестна как «провирус» и реплицируется как часть нормального клеточного генома. Регуляция экспрессии генов ВИЧвключает в себя комплексное взаимодействие между хроматин-ассоциированной провирусной ДНК, клеточным фактором транскрипции и вирусным трансактиватором транскрипции Tat [34]. Несплайсированная полноразмерная РНК обладает характеристиками эукариотической клеточной мРНК, поскольку содержит кэп на 5’-конце и полиаденилирована на 3’-конце [35]. Данная РНК используется как для продукции вирусных структурных и регуляторных белков, так и для формирования новых вирусных частиц.
Транскрипция провирусной ДНК и синтез вирусных белков
LTR ВИЧ служат сайтами инициации транскрипции и содержат cis-актиновые элементы, необходимые для синтеза РНК. Транскрипция инициируется в области перехода U3/R LTR ВИЧ. Сайт U3 содержит множество элементов (LBP-1, NFκB, LEF, Ets, USF-1, NFAT-1), которые направляют РНК-полимеразу II (pol II) к провирусной ДНК ВИЧ. Транскрипция с LTR ВИЧприводит к синтезу большого количества (более 30) копий вирусной РНК [36].
Эти РНК разделяются на 3 больших класса:
. несплайсированные РНК, которые функционируют как мРНК для Gag и прекурсора полипротеина Gag-Pol, и данные РНК пакуются в дочерние вирионы как генетический материал нового вируса;
. частично сплайсированные мРНК размером ~5 т.п.н., кодирующие вирусные белки Env, Vpu, Vif и Vpr;
. небольшие, размером около 1,7-2,0 т.п.н., полностью сплайсированные мРНК, которые транслируются в белки Rev, Tat и Nef.
На первой стадии ВИЧ-инфекции образуются полностью сплайсированные небольшие мРНК, инициирующие транскрипцию ранних регуляторных белков ВИЧTat, Rev и Nef [37-39]. Белок Tat связывается с TAR-петлей, формирующейся на 5’-конце продукта транскрипции и значительно ускоряет транскрипцию [34, 40]. Белок Rev взаимодействует с участком РНК RRE (Rev Response Element), образующимся на поздней фазе репликации ВИЧ. Данный комплекс из RRE и нескольких молекул Rev экспортируется в цитоплазму с помощью Rev-зависимого пути экспорта. Для этого комплекс Rev-RRE использует комплекс человеческих белков, содержащих белок экспортин-1 (XPO1/CRM1) и Ran-GTP. Образовавшийся комплекс Rev-RRE-Xpo1/RanGTP уже экспортируется в цитоплазму клетки и транслируется для синтеза остальных белков ВИЧили пакуется в качестве генома новых копий вируса [41].
Обычно трансляция мРНК в эукариотических клетках включает в себя поиск стартового кодона AUG, однако поскольку LTR ВИЧсодержат множество структурных регионов, таких как TAR, PBS, поли(А)-хвост, то обычный рибосомальный механизм поиска последовательности AUG на мРНК ВИЧне эффективен[42]. Кроме того, некоторые мРНК ВИЧсодержат последовательность AUG в области UTR (untranslated region), что может помешать инициации трансляции с истинного стартового кодона. Было предложено два основныхмеханизма, позволяющих инициировать трансляцию мРНК ВИЧальтернативными способами. Поскольку мРНК полиаденилирована, то она может быть транслирована через обычную кэп-зависимую инициацию трансляции или через связывание IRES (англ. Internal Ribosome Entry Site - участок внутренней посадки рибосомы[43-45]. После связывания с рибосомой РНК становится матрицей для синтеза вирусных белков.
Сборка вириона и продукция вируса
После синтеза полного набора вирусных белков начинается процесс сборки нового вириона ВИЧ. Для сборки инфективного вириона необходимо наличие двух копий вирусной РНК, вирусного праймера обратной транскрипции tRNALys3[26], полипротеинов Env, Gag и трех вирусных ферментов: протеазы, обратной транскриптазы и интегразы. Сборка ВИЧпроисходит на плазматической мембране инфицированной клетки. Все компоненты, необходимые для сборки вируса, доставляются к плазматической мембране с помощью транспортных систем инфицированной клетки [46].
Главную роль в сборке вируса играет прекурсор полипротеин Gag, Pr55Gag[47, 48]. Он состоит из матрикса (МА), капсида (CA), нуклеокапсида (NC), домена p6, а также включает два белка SP1 и SP2.МА-домен, ответственный за связывание Pr55Gag с клеточной мембраной, способствует включению вирусных гликопротеинов Env в образующиеся вирионы ВИЧ. СА обеспечивает мультимеризацию Gag в процессе сборки вириона, NC присоединяет вирусные РНК и способствует сборке вириона, домен p6 присоединяет ESCRT (the endosomal sorting complexes required for transport), который катализирует процесс отделения от мембраны клетки, способствуя, таким образом, отпочкованию вируса [49, 50].
.5 Механизмы рекомбинации ВИЧ
Скорость рекомбинации ретровирусов выше, чем для большинства других вирусов, а скорость рекомбинации ВИЧ выше, чем у других ретровирусов, таких как вирус лейкемии мышей и вирус некроза селезенки[51-54]. Анализ геномов ВИЧ путем прямого секвенирования показал, что рекомбинации в процессе синтеза ДНК - явление довольно частое [55-59]. Во время синтеза (-)цепи ДНК ОТ ВИЧможет «переключаться» между двумя РНК, упакованными в вирион, с использованием каждой из них в качестве матрицы для синтеза ДНК. Таким образом, рекомбинантную ДНК могут генерировать только те вирионы, в которых были упакованы две генетически различные молекулы РНК [60].
Необходимо соблюдение нескольких условий для генерации новой рекомбинантной формы ВИЧ: во-первых, клетка, в которой происходит репликация вируса, должна быть инфицирована более чем одним вирусом. Во-вторых, молекулы РНК от двух разных провирусов должны быть упакованы в один вирион. В третьих, во время обратной транскрипции ОТ должна переключаться между матрицами РНК для генерирования рекомбинантной ДНК-копии ВИЧ, которая должна быть интегрирована в геном клетки-мишени. И, наконец, такой рекомбинантный провирус должен обладать свойством генерировать вирусы, способные к инфицированию и репликации. По этим причинам факторы, влияющие на любой из таких этапов, могут оказывать влияние на рекомбинацию. В настоящее время мало известно о том, как часто клетки-мишени у пациентов инфицируются более чем одним ВИЧ-1 (двойная инфекция). В культуре клеток двойная инфекция встречается чаще, чем предполагалось, в сравнении со случайными событиями [61]. Такой результат, по крайней мере, связан с тем, что некоторые клетки имеют больше рецепторов/корецепторов и, следовательно, более восприимчивы к инфекции ВИЧ-1 [62]. Двойная инфекция возрастает, когда ВИЧ-1 передается через клеточно-опосредованные события, поскольку множество вирусов передаются от клетки-донора в клетки-мишени [61].
Во время сборки ВИЧ-1белок p24 упаковывает полноразмерную РНК в виде диметра, следовательно, выбор молекулы РНК происходит до капсидирования РНК-геномов. Основным фактором, определяющим выбор той или иной РНК, является сигнал инициации димеризации (dimerization initiation signal, DIS), расположенный в 5’-области нетранслируемой РНК ВИЧ-1 [63]. Большинство ВИЧ-1 субтипов B и D имеют последовательность GCGCGC в DIS, тогда как большинство ВИЧ-1 субтипов А, С, F и G имеют последовательность GTGCAC, хотя возможны и другие последовательности [64,65]. Считается, что палиндромная последовательность в DIS способствует связыванию оснований двух РНК, что инициирует димеризацию РНК. Другие последовательности нуклеотидов в геноме вируса также могут оказывать влияние на частоту РНК гетеродимеризации, хотя и с менее выраженным эффектом, чем DIS [66].
Скорость рекомбинации ВИЧ-1 была измерена с использованием маркерных генов. Эти результаты показали, что скорость рекомбинации пропорциональна увеличению расстояния между двумя аллелями, когда расстояние между маркерами составляет менее 600 нуклеотидов, а максимально возможная скорость рекомбинации достигается, когда две аллели разделены между собой 1300 нуклеотидами [54, 67].
Несмотря на то, что рекомбинация происходит на протяжении всего генома ВИЧ-1, структура РНК может влиять на частоту рекомбинации определенных областей[68]. Гомология нуклеотидных последовательностей может влиять как на скорость рекомбинации ВИЧ, так и на распределение точек разрыва генома [69, 70]. Например, рекомбинация возникает гораздо чаще между двумя РНК ВИЧ-1 одного субтипа, чем между РНК различных субтипов, упакованных вместе в один вирион [59].
Существует несколько предположений о причинах рекомбинации генетического материала ретровирусов. Одна из теорий гласит, что адаптивное значение рекомбинации происходит от ее способности восстанавливать генетические повреждения, в том числеу РНК-содержащих вирусов[71]. Действительно, в первых работах по изучению рекомбинации ретровирусов было высказано предположение о существовании модели «принудительного выбора» (forced copy-choice model), когда ОТ переключается с одной матрицы РНК на другую, если в матрице имеются повреждения (разрывы) [72]. Однако дальнейшие исследования показали, что репликация вируса происходит с одинаковой частотой как на целых, так и на поврежденных матрицах РНК [73], и экспериментально индуцированные генетические нарушения в РНК не привели к более высокой частоте возникновения рекомбинаций [74]. В дальнейшем эта модель была пересмотрена, дополнена и переименована в модель «динамического выбора» (dynamic copy-choice model) [75]. Эта модель предполагает, что баланс между полимеразной активностью и активностью РНКазы Н вирусной ОТ определяет стабильность связи между синтезируемой ДНК и матрицей РНК, и что смещение этого баланса влияет на скорость рекомбинации. Если баланс смещается в сторону большей активности РНКазы Н по отношению к полимеразной активности ОТ, то возникает снижение взаимодействия между синтезируемой ДНК и матрицей РНК из-за диссоциации гетеродуплекса РНК/ДНК и происходит переключение ОТ на другую РНК-матрицу [75]. Обратный эффект, при котором активность РНКазы H снижается, а полимеразная активность ОТ возрастает, показал снижение частоты переключения между матрицами РНК [76, 77].