Материал: Кузнецова Л.В., Бабаджан В.Д., Харченко Н.В. та ін. Імунологія

16 |
|
ІМУНОЛОГІЯ |
||
|
|
|
Продовження таблиці |
|
|
|
|
|
|
1 |
2 |
3 |
4 |
|
CD38 |
-- |
T- і B-лімфоцити, ак- |
Присутній на T-лімфоцитах кіркової |
|
|
|
тивовані В-лімфоцити |
речовини тимусу, активованих T-лім- |
|
|
|
|
фоцитах, незрілих B-лімфоцитах і |
|
|
|
|
плазматичних клітинах, бере участь |
|
|
|
|
в регуляції функцій B-лімфоцитів |
|
|
|
|
|
|
CD43 |
ICAM-1 |
T- і B-лімфоцити, гра- |
Бере участь в активації T-лімфоцитів |
|
|
|
нулоцити, моноцити |
|
|
|
|
|
|
|
CD45 |
-- |
Всі лейкоцити |
Бере участь в активації лімфоцитів, |
|
|
|
|
внутрішньоклітинна частина рецеп- |
|
|
|
|
тора є тирозинкіназою |
|
|
|
|
|
|
CD45RO |
-- |
T- і B-лімфоцити, гра- |
Маркер клітин пам'яті (лімфоцитів |
|
|
|
нулоцити, моноцити |
CD4), ефекторних клітин |
|
|
|
|
|
|
CD45RA |
-- |
Всі лейкоцити |
Маркер Т і В-лімфоцитів та моноци- |
|
|
|
|
тів |
|
|
|
|
|
|
CD45RB |
-- |
Т- і В– лімфоцити, мо- |
Маркер зрілих Т і В-лімфоцитів та |
|
|
|
ноцити, гранулоцити |
моноцитів |
|
|
|
|
|
|
CD71 |
Трансферин |
T-лімфоцити, моно- |
Рецептор трансферину, маркер акти- |
|
|
|
цити |
вованих T-лімфоцитів |
|
|
|
Маркери B-лімфоцитів |
|
|
Поверхневі |
Антиген |
B-лімфоцити |
Присутні тільки на зрілих B-лімфо- |
|
імуногло- |
|
|
цитах |
|
буліни |
|
|
|
|
|
|
|
|
|
CD10 |
-- |
B-лімфоцити |
Присутній на незрілих B-лімфоци- |
|
|
|
|
тах, з'являється на лейкозних клітках |
|
|
|
|
при гострому лімфолейкозі |
|
|
|
|
|
|
CD19 |
-- |
B-лімфоцити |
Присутній на пре-B-лімфоцитах і на |
|
|
|
|
всіх зрілих B-лімфоцитах, бере |
|
|
|
|
участь в активації B-лімфоцитів |
|
|
|
|
|
|
CD20 |
-- |
B-лімфоцити |
Присутній на всіх B-лімфоцитах |
|
|
|
|
|
|
CD21 |
C3d, CD23 |
B-лімфоцити |
Рецептор до комплементу і вірусу |
|
|
|
|
Епштейн-Бара |
|
|
|
|
|
|
CD23 |
IgE |
B- і T-лімфоцити, мо- |
Низькоафінний рецептор до Fc-фраг- |
|
|
|
ноцити, еозинофіли |
менту IgE |
|
|
|
|
|
|
CD32 |
IgG |
B-лімфоцити, грану- |
Нізькоафінний рецептор до Fc-фраг- |
|
|
|
лоцити |
менту IgG |
|
|
|
|
|
|
CD40 |
gp39 |
B-лімфоцити |
Стимулює проліферацію B-лімфоци- |
|
|
|
|
тів, за будовою схожий з CD27 і ре- |
|
|
|
|
цептором чинника некрозу пухлин |
|
CD72 |
CD5 |
B-лімфоцити |
З’являється на кістково-мозкових по- |
|
|
|
|
передниках B-лімфоцитів, бере |
|
|
|
|
участь в їх диференціюванні |
|
|
|
|
|
|
|
ПРИНЦИПИ ФУНКЦІОНУВАННЯ ІМУННОЇ СИСТЕМИ |
17 |
|||
|
|
|
|
|
|
|
|
|
Продовження таблиці |
||
|
|
|
|
|
|
1 |
2 |
3 |
4 |
|
|
HLA-DR |
Антиген, |
B- і T-лімфоцити, мо- |
Антиген MHC II класу, бере участь в |
|
|
|
CD4 |
ноцити |
представленні антигену T-хелперам і |
|
|
|
|
|
їх активації, маркер активованих T- |
|
|
|
|
|
лімфоцитів |
|
|
|
|
Маркери моноцитів і |
макрофагів |
|
|
CD11a |
ICAM-1 |
Всі лейкоцити |
Альфа-ланцюг LFA-1, бере участь в |
|
|
|
|
|
міжклітинній адгезії |
|
|
|
|
|
|
|
|
CD11b |
C3bi, фібро- |
Моноцити, грануло- |
Альфа-ланцюг CR3, бере участь в |
|
|
|
нектин |
цити, NK-лімфоцити |
міжклітинній адгезії |
|
|
CD11c |
C3bi |
Моноцити, грануло- |
Альфа-ланцюг CR4, бере участь в |
|
|
|
|
цити, B- і NK-лімфо- |
міжклітинній адгезії |
|
|
|
|
цити |
|
|
|
CD14 |
TLR4 |
Моноцити, макрофаги |
Приймає участь в активації цих клі- |
|
|
|
|
|
тин через Toll-подібний рецептор-4 |
|
|
|
|
|
|
|
|
CD18 |
-- |
Всі лейкоцити |
Бета-ланцюг рецепторів |
|
|
|
|
|
CD11a/CD18, CD11b/CD18, |
|
|
|
|
|
CD11c/CD18, бере участь в міжклі- |
|
|
|
|
|
тинній адгезії |
|
|
|
|
|
|
|
|
|
|
Маркери NK-лімфоцитів |
|
|
|
CD3 |
-- |
Зрілі NK- і Т-лімфо- |
|
|
|
|
|
цити |
|
|
|
|
|
|
|
|
|
CD16 |
Fc-фрагмент |
NK-лімфоцити, моно- |
Низькоафінний рецептор IGG |
|
|
|
IgG |
цити і гранулоцити |
|
|
|
|
|
|
|
|
|
CD56 |
-- |
NK- і T-лімфоцити |
Присутній на частині T-лімфоцитів, |
|
|
|
|
|
бере участь в міжклітинній адгезії, |
|
|
|
|
|
NCAM (neural cell adhesion molecule) |
|
|
|
|
|
|
|
|
CD57 |
-- |
NK- і T-лімфоцити |
Присутній на частині лімфоцитів |
|
|
|
|
|
CD8, при деяких вірусних інфекціях |
|
|
|
|
|
збільшується число лімфоцитів, що |
|
|
|
|
|
несуть одночасно CD8 і CD57 |
|
|
|
|
|
|
|
|
Примітка: (–) - невідомий або відсутній; CR - рецептор до компонентів комплементу; ICAM - молекули міжклітинної адгезії (Inter Cellular Adhesion Molecule); LFA - лімфоцитарний функціональний антиген (Lymphocyte Function-associated Antigen).
Лімфоцити - це єдині клітини організму, здатні специфічно розпізнавати і розрізняти різні антигени і відповідати активацією на контакт з певним антигеном. При схожій морфології малі лімфоцити діляться на дві популяції, що мають різні функції і продукують різні білки.

18 |
ІМУНОЛОГІЯ |
В-лімфоцити. Одна з популяцій лімфоцитів отримала назву В-лім- фоцити, від назви органу "бурса Фабріциуса", де було вперше виявлено дозрівання цих клітин у птахів. Маркер CD20 представлений на В-клі- тинах всіх стадій розвитку В-лімфоцитів. У людини В-лімфоцити дозрівають в кістковому мозку. В-лімфоцити розпізнають антигени специфічними рецепторами імуноглобулінової природи (CD19-22), які у міру дозрівання експресуються на їх мембранах. Взаємодія антигену з такими рецепторами є сигналом активації В-лімфоцитів, та їх антигензалежного диференціювання в плазматичні клітини, що активно продукують і секретують специфічні для даного антигену антитіла - імуноглобуліни.
При дозріванні В-лімфоцити змінюють клас імуноглобулінів, які синтезуються ними. Спочатку В-лімфоцити синтезують імуноглобуліни класу М (IgM), при дозріванні 10 % В-лімфоцитів продовжують синтезувати IgM, 70 % перемикаються на синтез IgG, а 20 % - на синтез IgA. Наступна експресія поверхневого IgD означає, що клітина готова до стимуляції антигеном. Деякі клітини, таким чином, несуть поверхневі Ig трьох різних класів: M, G і D або M, A і D, але усі молекули Ig на одній клітині мають однаковий ідіотип і, отже, кодуються одними і тими ж генами V(H) і V(L). Після стимуляції антигеном поверхневий IgD втрачається і у клітин пам'яті не виявляється. Антигенна структура зрілого В-лімфоциту: CD19, CD20, CD21, CD45R, CD40, MHC II кл., IgM, IgD. Окрім молекул рецепторного комплексу на поверхні В-клітин експресуються молекули гістосумісності MHC II кл., оскільки В-лімфо- цити є антиген-презентуючими клітинами.
В-лімфоцити складаються з декількох субпопуляцій:
1)В1 - лімфоцити - попередники плазмоцитів, несуть на мембрані диференційований антиген CD5+, синтезують антитіла IgM після контакту з антигеном без взаємодії з Т-лімфоцитами;
2)В2 - лімфоцити - попередники плазмоцитів, проходять диференціювання в кістковому мозку від стовбурової клітини до попередників В- лімфоцитів під впливом ростових чинників, інтерлейкінів (IL-1, 4, 6) синтезують імуноглобуліни усіх класів після контакту з антигеном у відповідь на взаємодію з Т-хелперами. Ці клітини забезпечують гуморальний імунітет на антигени, розпізнавані Т-хелперами;
3)В3-лімфоцити (К-клітини), або В-кілери, вбивають клітини-анти- гени, покриті антитілами. Відносяться до великих гранулярних лімфоцитів, здатних розпізнавати (як і T-клітини ) зміни клітинної поверхні, які виникають при злоякісному переродженні чи вірусній інфекції. Крім
ПРИНЦИПИ ФУНКЦІОНУВАННЯ ІМУННОЇ СИСТЕМИ |
19 |
|
|
того, на відміну від цитотоксичних T-лімфоцитів, вони ефективно розпізнають клітини, поверхня яких не має молекул MHC або частково їх втратила;
4) В-супресори гальмують функцію Т-хелперів, а В-лімфоцити пам'яті, зберігаючи і передаючи пам'ять про антигени, активно синтезують певні імуноглобуліни при повторній зустрічі з антигеном.
Особливістю В-лімфоцитів є те, що вони спеціалізуються на конкретних антигенах. При реакції В-лімфоцитів з антигеном, що зустрічається уперше, утворюються плазмоцити, які виділяють антитіла саме проти цього антигену. Утворюються клони В-лімфоцитів, відповідальні за реакцію з цим конкретним антигеном. При повторній реакції розмножуються і синтезують антитіла тільки В-лімфоцити, а точніше - плазмоцити, спрямовані проти цього антигену. Інші клони В-лімфоцитів не беруть участь в реакції. В-лімфоцити безпосередньо не приймають участь в боротьбі з антигенами. Під впливом стимулів від фагоцитів і Т- хелперів вони трансформуються в плазмоцити, які і синтезують антитіла імуноглобуліни, які знешкоджують антигени.
Т-лімфоцити отримали свою назву у зв'язку з їх диференціюванням у тимусі. Зрілі Т-лімфоцити (CD2, CD3) на відміну від незрілих (тимоцитів – CD2) здатні відповідати проліферацією на Т-клітинні мітогени. За функціями серед Т-лімфоцитів розрізняють ефекторні (CD8 цитотоксичні лімфоцити – CTL, T-кілери) і регуляторні (CD4+ Т-хелпери-Th) субпопуляції.
Т-хелпери стимулюють проліферацію і диференціювання цитотоксичних лімфоцитів, В-клітин і утворення антитіл. Тобто, Т-хелпери мають хелперну функцію (стимулюють В-лімфоцити для продукції імуноглобулінів) і індукторну функцію (стимулюють проліферацію і диференціювання цитотоксичних лімфоцитів, що відповідають на розчинні антигени проліферацією і продукцією лімфокінів).
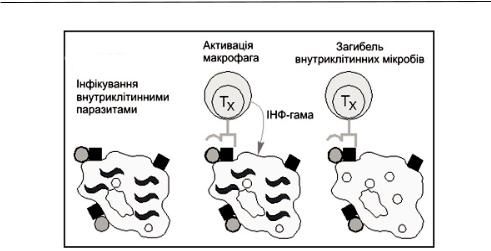
Внутрішньоклітинні паразити, здатні вижити усередині макрофагів, руйнують механізми знищення, властиві цим клітинам. Проте такі мікроорганізми не здатні перешкодити макрофагам переробити невеликі фрагменти антигенів (неповний фагоцитоз) і експонувати їх на своїй поверхні (процесінг). T–хелпери, відзначені цими антигенами, здатні впізнавати комбінацію з антигену і молекули MHC II класу на поверхні макрофага і зв'язуватися з нею (антигенпредставлення), а потім продукувати цитокіни (інтерферон-γ), які активують макрофаги, запускаючи пошкоджені раніше мікробіцидні механізми макрофагів, і викликають загибель внутрішньоклітинних мікроорганізмів – повний фагоцитоз (рис. 1).
20 |
ІМУНОЛОГІЯ |
Рис. 1. Активація макрофагів Т - хелперами
Примітка: кружки - поверхневий мікробний антиген; квадрати - молекули MHC класу II, хвилясті лінії - внутрішньоклітинні паразити.
Існує дві субпопуляції CD4+ Т-хелперів - Т-хелпери 1 і 2 типів, що не мають відмінностей за антигенною структурою, але розрізняються за набором (профілем) цитокінів, які вони здатні синтезувати у відповідь на антигенну стимуляцію, і від цього профілю залежить, який з двох основних типів імунної відповіді буде реалізований (клітинний або гуморальний).
Т-хелпери 1 типу (Th1) мають диференційні антигени CD3, CD4, CD29, CD45Ra. Це - активатори клітинного імунітету, натуральних кілерів і моноцитів. Якщо наївна Т-клітина розпізнає антиген, що презентується макрофагом, то вона трансформується в T-хелпер 1 типу. На цій клітині з’являються маркери диференціювання CD25 та CD45RB. Функція таких клітин - посилення активності макрофагів, спрямованої на знищення захопленого антигену, або приведення його в імуногенну форму.
Продукуючи інтерлейкіни-2, 3, 12, ІФН-γ і ФНО-β, ГМ-КСФ, вони викликають активацію цитотоксичних Т-лімфоцитів, натуральних кілерів, макрофагів та Т-ефектори гіперчутливості уповільненого типу. Thl забезпечують імунітет проти вірусів, внутріклітинних бактерій і онкогенних клітин. Активність Th1 подавляє інтерлейкін-10.
Т-хелпери 2 типу (Th2) мають диференційовочні антигени CD3, CD4, CD29, CD45Ra і відповідають за кооперацію з В-клітинами. Якщо Т-клітина розпізнає антиген, що розміщений на поверхні В-лімфоцитів, то це розпізнавання є сигналом до трансформації в Т-хелпери 2 типу, які