Материал: Жукова Л.А. (отв. ред.) Онтогенетический атлас лекарственных растений. Том VII
ная высота их равна 100,0 см и выше. Соцветия достигают 6,0 см в длину. Число колосков на соцветии насчитывается от 17 до 55. Корневая система состоит в среднем из 54,0 корней длиной 14,2 см. Высокая изменчивость наблюдается у таких признаков, как число вегетативных побегов (CV = 43,1 %), число колосков в соцветии (CV = 40,9 %), длина листа на генеративном побеге (CV = 64,1 %), число корней (CV = 36,8 %), а также размер дерновины (CV = 44,1 %).
СТАРЫЕ ГЕНЕРАТИВНЫЕ растения. Переход к старому генеративному состоянию выражается в увеличении числа отмерших побегов на дерновину. Размер дерновины в этом состоянии достигает 32,6 см в длину и 22,2 см в ширину. У старых генеративных растений в среднем насчитывается 14,0 (от 2 до 54 шт.) генеративных побегов, средняя высота – 91,3 см. Число вегетативных побегов от 45 до 480. Соцветие достигает в среднем 7,2 см в длину и состоит из 47,1 колосков. Корневая система представлена в среднем 42,2 придаточными корнями (максимальное число 285 шт.), длиной 17,8 см. Структура дерновины на в этом состоянии становится рыхлой, с отмершей частью в центре. Отмершая часть в дерновине составляет 20–30 %.
СУБСЕНИЛЬНЫЕ особи представлены дерновинами с отмершей центральной частью подковообразной формы. Генеративные побеги полностью отсутствуют. Размер дерновины в этом состоянии изменяется от 21 до 28 см в длину и от 24 до 34 см в ширину. Дерновина в целом состоит из 44,5 вегетативных побегов высотой 20,9 см. Всего листьев на дерновине насчитывается в среднем 81,3 шт. Отмершая (лысая) часть дерновины занимает в среднем 50–60 % от всей площади. Процессы отмирания преобладают над новообразованием.
СЕНИЛЬНОЕ онтогенетическое состояние характеризуется большим размером дерновины и малым числом побегов. Дерновина распадается на «полукольца». Живая часть дерновины может существовать на одной половине, а другая часть дерновины полностью отмирает. Размер дерновины в этом состоянии в среднем 18,6 см (длина) и 32,3 см (ширина). Дерновина состоит из 13,8 живых вегетативных побегов высотой 17,7 см. Отмершая часть дерновины занимает в среднем 70–80 % от всей площади.
Ломкоколосник дернистый относится к II надтипу онтогенеза (полный онтогенез осуществляется в ряду поколений вегетативно возникших особей), В-типу, т. е. онтогенез семенной особи несколько сокращен и завершается старческой партикуляцией в старом генеративном и субсенильном состояниях, партикулы при этом не омолаживаются
(Жукова, 1995).
256
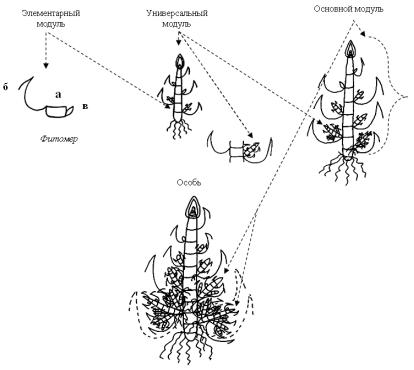
Проведено структурно-морфологическое исследование особей P. caespitosa в первый год жизни на базе модульного подхода. Использовали три категории модулей – элементарный (фитомер побега), универсальный (одноосный побег) и основной модуль (пространственновременная структура) (Савиных, 2002; 2007). Элементарный модуль включает короткое междоузлие, узел с листом на его верхнем конце и боковую почку возобновления, расположенную на нижнем конце междоузлия супротивно листу (рис. 56).
Рис. 56. Модульная организация ломкоколосника дернистого:
а – узел; б – лист; в – почка
Изучение модульной структуры ломкоколосника дернистого в условиях интродукции показало, что универсальный модуль в 1-й год жизни представлен укороченным (розеточным) вегетативным побегом. Универсальные модули любого порядка образуют собственные корни на базальной части и приступает к кущению, формируя собственную зону
257
кущения, при развертывании своего 4–5-го листа. Тип развития побегов внутривлагалищный. На основецелого универсального модуля или его части формируется модуль высшего ранга – основной модуль. У Psathyrostachys caespitosa на 1-ом году жизни основные модули образуются на основе побегов II порядка и включают побеги III–IV порядков. Они, имея собственную корневую систему, функционируют как целостная система. Между главным побегом и боковыми побегами II–IV порядков, а также между побегами разных порядков существует тесная взаимосвязь, влияющая на их развитие (Скобелева, 2011).
В целом модель побегообразования ломкоколосника дернистого представлена в виде полурозеточной симподиальной системы побега кущения. Побег как универсальный модуль может быть или розеточным вегетативным, или полурозеточным монокарпическим, ди-, полициклическим генеративным (Модульная организация..., 2011).
Большинство изученных ценопопуляций Psathyrostachys caespitosa являются неполночленными, т. к. проростки были обнаружены единично и не во всех ценопопуляциях Выявлено 4 типа онтогенетического спектра: бимодальный, левосторонний, правосторонний и центрированный. Из рассмотренных 18 ценопопуляций с левосторонним спектром было отмечено 38,9 %, с бимодальным спектром – 33,3 %, с правосторонним – 16,7 %, с центрированным – 11,1 %. Для P. caespitosa в данном диапазоне условий базовым спектром является бимодальный спектр с абсолютным подъемом на старых ss и локальным – на молодых im-v. Этот тип характерен для плотнодерновинных злаков с семенным возобновлением (Скобелева, 2012). Изучение виталитетного состава показало, что в большинстве ценопопуляций преобладают особи промежуточного класса В, что говорит о хорошем состоянии особей P. caespitosa в исследованных ценопопуляциях. Анализ виталитетной структуры показал, что структура ценопопуляций представлена 3 типами: преобладающей процветающей (69,2 %), депрессивной (23,1 %) и равновесной (7,7 %) (Злобин, 1989). Результат анализа оценки жизненности ценопопуляций P. caespitosa по размерному спектру особей показал, что в наиболее благоприятных условиях находятся растения в асс. Psathyrostachetum junceae. Индекс виталитета (IVC) высокий от 1,27 до 1,38. Также благоприятные условия отмечены в асс. Agropyretum cristati (IVC от 1,04 до 1,14). В ценопопуляциях с высокими показателями IVC сумма баллов по организменным и популяционным признакам высокая. В наименее благоприятных условиях (с низкими показателями суммы баллов от 21 до 29) находятся особи P. caespitosa, где IVC низкий от 0,61–0,86. Как правило, эти ценопопуляции относятся к депрессивным.
258
Для выживания и устойчивого развития P. caespitosa в процессе эволюции выработал комбинированный защитно-стрессовый тип онтогенетической стратегии. По системе оценки эколого-фитоценотических стратегий Л. Г. Раменского – Дж. Грайма (Миркин и др., 1999), характеризующей основные направления конкурентной стратегии растений на выживание при стрессе и нарушениях, P. caespitosa обладает смешанным типом жизненной стратегии с сочетанием черт виолентности и патиентности – конкурентно-стресс-толерантным (CS). В благоприятных условиях усиливается С-составляющая, и злак проявляет виолентность – С-стратегию. В условиях сильного стресса вид проявляет себя как S-стратег, т. е. патиент. Проявлением черт виолентности (С) является доминирование или содоминирование в степных сообществах. Черты виолентности проявляются и в том, что вид имеет крупный габитус, много побегов и относительно большую общую листовую поверхность. Показателями патиентности (S) являются засухоустойчивость и солевыносливость.
259
38. Онтогенез ковыля перистого
(Stipa pennata L.)
Ковыль перистый (Poaceae) – это опушечно-степной плотнодерновинный злак, травянистый многолетник, очень декоративный в фазе плодоношения, высотой 30–100 см, ксерофит, мезотроф, гелиофит (Флора ботанического сада.., 2012) с интравагинальными (внутривлагалищными) побегами. Пазушная почка в таком случае растет вертикально вверх внутри влагалища своего кроющего листа (Жизнь растений, 1984). Листья шириной 0,5–2,0 мм, свернутые вдоль, реже плоские, на верхушке короткозаостренные, у молодых листьев – с кисточкой из волосков длиной до 3 мм; язычок длиной 0,7–3 мм; влагалища голые или реснитчатые. Соцветие – метелка (Stipa pennata, http://www.plantarium.ru)
длиной 3,0–5,0 см, сжатое, из 6–20 колосков. Колоски беловатые; колосковые чешуи длиной 3–5 см, длиннозаостренные; цветковые чешуи длиной (15)16–17(21) мм, внизу сплошь, а выше с 7 рядами волосков, с остью длиной 25–35 см, ость коленчато-согнутая, в нижней скрученной части каштанового цвета голая, выше перистая, с волосками длиной около 5 мм (Stipa pennata, http://www.plantarium.ru). Цветет в апреле-
мае, плодоносит в мае-июне. Плоды – зерновки, ими размножается. Дает самосев. Устойчив в культуре
Европейско-среднесибирско-древнесредиземноморский вид. В России распространен на юге европейской части и на юге Сибири. Растет по степям, на остепненных лугах и полянах, по обнажениям мела и известняка. Сокращаются площади его естественного произрастания, поэтому растение занесено в Красную книгу России (статус 2), а также в региональные Красные книги: Белгородской (статус 2), Воронежской (статус 3), Курской (статус 3), Липецкой (статус 2) областей. Декоративное, кормовое и фитомелиоративное растение (Флора ботанического сада.., 2012).
Онтоморморфогенез ковыля перистого исследовали на примере особей, произрастающих в коллекции «Систематикум флоры Центрального Черноземья» и на экспозиции «Степи Центрального Черноземья». Семенной материал репродукции заповедника «Галичья Гора» высевался в закрытый грунт весной (конец марта). Изучение онтогенеза проведено согласно работам Т. А. Работнова (1950), А. А. Уранова (1967) и их учеников (Ценопопуляции..., 1976; 1988). Онтогенез ковыля представлен на рисунках 57–59.
ПЛОД – узкоцилиндрическая, заостренная зерновка, длиной 1,0– 1,2 см. Завязь одногнездная, с одним семязачатком (Жизнь растений.., 1984). Зародыш хорошо дифференцирован, с мощным эндоспермом (Николаева и др., 1985).
260