Материал: Жукова Л.А. (отв. ред.) Онтогенетический атлас лекарственных растений. Том VII
8. Онтогенез тимьяна Маршалла
(Thymus marschallianus Willd.)
Thymus marschallianus Willd. (тимьян Маршалла) представитель рода Thymus L. (сем. Lamiaceae). Жизненная форма вида – неподвижный полукустарничек с ветвящимися моно- и дициклическими генеративными побегами. Для вида характерны прямостоячие или приподнимающиеся вегетативные и генеративные побеги, высота последних может достигать 30 см. Листья продолговатые или ланцетные, тупо заостренные (длина 10–20 мм, ширина 5–6 мм), короткочерешковые или почти сидячие, голые. Соцветие от продолговато-яйцевидного до удлиненного, иногда с расставленными многоцветковыми мутовками; чашечки окрашенные, волосистые, зубчики по краю длиннореснитчатые (Доронькин, 1997).
T. marschallianus, как и остальные виды рода, активно используют в официальной и народной медицине (Верещагин и др., 1959). Источником лекарственного растительного сырья является надземная травянистая часть растения. Химические компоненты, входящие в состав растительного сырья вида, разнообразны и представлены алициклическими соединениями, моно- и сесквитерпеноидами, фенолкарбоновыми кислотами, флаваноидами, жирными кислотами и их производными и др. Активным веществом является эфирное масло, в надземной части растения его количество достигает 0,8 % (Растительные…, 2011).
Ареал T. marschallianus протягивается от Австрии и Чехии на западе до Алтая на востоке. Г. А. Пешковой (2001) вид отнесен к группе евро- пейско-западно-центрально-азиатских видов. Для T. marschallianus характерна приуроченность к степной и лесостепной зонам.
Для описания онтогенеза использован материал, собранный в злако- во-типчаковой настоящей мелкодерновинной степи в окрестностях оз. Кучукское Алтайского края.
Изучение онтогенеза проводили с использованием традиционных методов и подходов (Работнов, 1950; Уранов, 1975; Ценопопуляции…, 1976). Выборка в каждом онтогенетическом состоянии составила 50 особей.
ПЛОД T. marschallianus – ценобий. По данным Е. Е. Гогиной (1975), при созревании ценобия 4 эрема быстро отделяются. Созревание отдельного эрема происходит быстро, около четырех-пяти недель. Созревшие эремы некоторое время задерживаются в чашечке благодаря волосистому кольцу по ее зеву. Их высыпание происходит при резком сотрясении или раскачивании генеративного побега. При этом основная масса осыпавшихся эремов оказывается в непосредственной близости от
111
материнского растения. Прорастание семян в основном происходит
вконце вегетационного периода.
УПРОРОСТКОВ формируются почти округлые семядоли со слабо сердцевидным основанием, а также 2–3 пары настоящих листьев, без четко выраженного черешка. Побег проростка состоит из укороченных метамеров (зачастую 3-х), его рост завершается формированием зимующей терминальной почки. Высота побега достигает 0,3 см. Как правило, большинство проростков сохраняется на следующий год.
В ЮВЕНИЛЬНОЕ состояние растение переходит на второй год. На розеточной части первичного побега сохраняются остатки прошлогодних листьев. Они защищают терминальную почку во время неблагоприятных условий. Годичный побег нарастает моноподиально и является верхнерозеточным. На годичном побеге образуется 3–4 пары супротивных листьев. Главный корень начинает ветвиться. На нем образуется несколько нитевидных боковых корней. К концу вегетационного периода первичный побег почти полностью полегает, что обеспечивает его сохранение в зимний период.
На третий год у особей в ИММАТУРНОМ онтогенетическом состоянии из терминальной почки развивается верхнерозеточный годичный побег. Таким образом, структура первичного побега представлена 3-мя годичными приростами: первый – розеточный, второй и третий – верхнерозеточные. Такой розеточно-верхнерозеточный (Колегова, Черемушкина, 2012) первичный побег имеет восходящее положение. Первые два годичных прироста расположены плагиотропно и к этому времени уже одревесневают, а третий имеет ортотропное положение и остается полностью травянистым. Последний годичный побег более мощный, его длина равна или превышает общую длину прошлогодних. Он ветвится в апикальной и средней частях. Ветвление происходит не только на третьем, но и на первом годичном приросте. В его узлах к концу вегетационного периода из спящих почек развиваются от 2 до 6 розеточных побегов. Также хорошо заметны спящие почки в узле базального междоузлия второго годичного прироста. Почки открытые, и за счет плагиотропного положения прироста они, как правило, сохраняются в зимний период. Корневая система также состоит из главного и боковых корней. Особи в имматурном состоянии представляют собой разветвленный первичный побег.
В зимний период отмирает весь прирост этого года. Базальная часть первичного побега, как правило, сохраняется и представлена первым годичным розеточным приростом и 1–2 удлиненными метамерами второго. Сохраняются также розеточные побеги в узлах первого годичного
112
прироста и иногда спящие почки в узле базального междоузлия второго верхнерозеточного прироста.
Вначале вегетационного периода в ВИРГИНИЛЬНОМ онтогенетическом состоянии все почки трогаются в рост. Растение представляет собой первичный куст. В базальной части куста развиваются озимые розе- точно-верхнерозеточные побеги. Розеточная часть озимых побегов сформировалась в узлах первого годичного прироста первичного побега прошлой осенью, остальная часть побега – верхнерозеточная, является годичным побегом. Как правило, озимые побеги имеют восходящее положение. При этом базальный участок – розеточный прирост – расположен плагиотропно и одревесневает полностью, а остальная часть побега располагается ортотропно и остается травянистой. Для розеточноверхнерозеточных побегов характерно ветвление. Так, в узлах базальной розеточной части из пролептических почек развиваются розеточные побеги ветвления. Из силлептических почек в средней части побега развиваются безрозеточные (или розеточные) побеги обогащения.
Вструктуре куста имеются еще и однолетние верхнерозеточные побеги. Образуются они из спящих почек в сохранившихся узлах второго годичного прироста первичного побега. Верхнерозеточные побеги травянистые, в связи с этим полностью отмирают в зимний период.
Озимые розеточно-верхнерозеточные побеги являются побегами ветвления (ПВ) n + 1 порядка. Высота побегов ветвления в среднем составляет 5,3 ± 0,7 см. Их одревесневшие базальные части впоследствии входят в многолетнюю основу куста и способствуют его разрастанию. Стоит отметить, что разрастание куста незначительное в связи с тем, что длина базальных участков побегов ветвления не превышает 1 см. Диаметр куста в среднем составляет 5,1 ± 0,1 см. В базальной части ПВ формируются многочисленные придаточные корни. Корневая система
уособей в виргинильном онтогенетическом состоянии смешанная и состоит из системы главного и придаточных корней.
ВМОЛОДОЕ ГЕНЕРАТИВНОЕ состояние растение переходит на следующий год. Фаза первичного куста сохраняется. В базальной части куста на сохранившихся многолетних участках ПВ n + 1 порядка развиваются ПВ n + 2 порядка. Часть ПВ n + 2 порядка переходит к цветению, образуются озимые генеративные побеги, а остальная часть остается вегетативной. Структура ПВ n + 2 порядка и место их образования соответствует структуре и локализации ПВ n + 1 порядка. Базальные части тех и других образовались прошлой осенью и представляют собой розеточный побег. Годичный генеративный побег по структуре безрозеточный. В средней и апикальной частях годичного побега происходит ветвление. Из силлептических почек в апикальной части развиваются
113
безрозеточные генеративные побеги, а в средней – безрозеточные вегетативные побеги обогащения. Последние могут ветвиться.
По структуре озимый генеративный побег полурозеточный. В базальной части озимых генеративных побегов к концу вегетационного периода, так же как и у озимых вегетативных побегов, из пролептических почек развиваются розеточные побеги, а в узле первого удлиненного междоузлия закладываются спящие почки. Кроме озимых генеративных побегов, в структуре куста обнаружены и моноциклические генеративные побеги, которые развиваются из спящих почек и представлены удлиненными метамерами. Для моноциклических генеративных побегов характерно акро- и мезотонное ветвление, аналогичное озимым генеративным побегам. Число генеративных побегов на растении не превышает 16, вегетативных побегов – 17. Куст незначительно разрастается, его диаметр в среднем 8,9 ± 0,1 см. Число отмерших побегов на растении может достигать 9. В зимний период вновь происходит отмирание побегов. Отмирает большая часть генеративного и вегетативного побегов.
Для генеративных побегов T. marschallianus всех типов характерна синфлоресценция. Она образована главным соцветием и 2–4 паракладиями I порядка. Соцветие, заканчивающее главный побег, и паракладии – кистевидный тирс, состоящий из мутовчато расположенных, расставленных дихазиев.
Вмолодом генеративном состоянии осуществляется семенное размножение. Данный способ у растений основной. Вегетативного размножения, несмотря на обилие придаточных корней, не происходит.
ВСРЕДНЕВОЗРАСТНОМ ГЕНЕРАТИВНОМ состоянии растение пред-
ставляет собой клон. Формирование клона происходит следующим образом. В базальной части первичного куста накапливаются многолетние части ПВ, куст разрыхляется. Начинается отделение укоренившейся системы ПВ. В результате образуется клон, состоящий из первичного куста и нескольких (часто 3) близко расположенных дочерних особей (кустящихся партикул). Первичный куст характеризуется наличием главного корня и первичного побега с 1–2 сохранившимися метамерами. Дочерние особи имеют сохранившиеся базальные остатки ПВ и придаточную корневую систему. Как правило, раметы находятся в том же онтогенетическом состоянии, что и материнская особь. Диаметр такого дезинтегрированного куста в среднем 22,1 ± 0,8 см. Число вегетативных побегов достигает 37, генеративных – 26. В средневозрастном генеративном состоянии клон может находиться несколько лет (3–5).
114
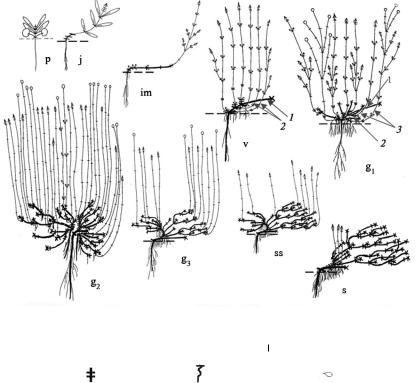
Рис. 12. Онтогенез тимьяна Маршалла (Thymus marschallianus Willd.)
Условные обозначения: ↑ – вегетативный побег;  – генеративный побег,
– генеративный побег,
×– отмерший побег; ▌ – одревесневший участок; │ – неодревесневший участок;
┼– узел; – сближенные узлы; – придаточный корень; – почка;
–– – – уровень почвы (у g2 уровень почвы не показан).
1 – одревесневший участок первичного побега; 2 – ПВ n + 1 порядка; 3 – ПВ n + 2 порядка; в g3–ss представлен первичный куст клона
ВСТАРОМ ГЕНЕРАТИВНОМ состоянии происходит накопление отмерших частей ПВ в структуре каждой партикулы клона. Новые побеги развиваются в основном по периферии кустов. Центральная часть клона все больше разрыхляется. Одна из партикул может полностью отмереть: кустящиеся партикулы в компактном клоне угнетающе действуют друг на друга. В связи с этим диаметр особи уменьшается. Среднее значение этого показателя составляет 10,3 ± 0,2 см. Число генеративных побегов достигает 15, вегетативных – 26. В главном корне начинаются процессы разрушения.
Особи клона в СУБСЕНИЛЬНОМ и СЕНИЛЬНОМ состояниях характеризуются увеличением числа остатков сухих ПВ как по периферии кустов, так и в их базальной части. Число отмерших партикул увеличи-
115