Курсовая работа (т): Генетика и биохимия алкоголизма
ГАМК - наиболее распространенный тормозный нейромедиатор в нервной системе млекопитающих. Связываясь со специфическими рецепторами на поверхности клеточной мембраны, он изменяет активность нейронов и полностью подавляет их восприимчивость к сигналам. Один класс таких рецепторов, ГАМКА, состоит из белковых субъединиц, расположенных вокруг ионного канала, по которому ионы хлора поступают в клетку. Изменения в GABRA2-гене, который кодирует одну из субъединиц рецептора ГАМКА, существенно влияет на бета-волны ЭЭГ, соответствующие нейронному растормаживанию.
Нейроны, несущие ГАМК-рецепторы, особенно многочисленны в лобных областях коры головного мозга. Полная утрата тормозной активности в этих участках может привести к эпилептическим судорогам, при которых обычно применяют препараты, повышающие активность ГАМК и вызывающие торможение. Частичная же утрата тормозной активности, опосредуемой ГАМК, по-видимому, приводит к потере самоконтроля или к импульсивности. Названные признаки составляют часть симптомокомплекса, характерного для ряда психических заболеваний, в число которых входят маниакально-депрессивный психоз, токсикомания, асоциальная психопатия.
Как показали исследования, нарушения в гене GABRA2, не сопровождающиеся изменением архитектуры рецептора ГАМКА, связаны с алкоголизмом. Вероятно, эти нарушения приводят к образованию дефектных субъединиц, что в свою очередь уменьшает число нормально функционирующих рецепторов.
Всего на сегодняшний день найдены десятки генов, нарушение работы которых, как предполагают, повышает риск развития алкоголизма.
Знание таких генов дает нам новую информацию о природе болезни, в данном случае алкоголизма. Открываются возможности для создания новых лекарств, ведь мишени для алкоголя могут быть в той же степени мишенями и для лекарственных веществ. Наконец, можно будет предсказывать риски развития алкоголизма у людей-носителей каких-либо «неблагополучных» генов. Но наследственная предрасположенность к заболеванию проявляется в том случае, если и гены, и среда действуют в одну сторону. Пока мы не можем менять гены, но мы можем изменить условия среды, чтобы снизить риск.
генетический
нейропсихический метаболизм алкоголизм
2. Биохимия алкоголизма
2.1 Действие алкоголя на мозг
.1.1 Молекулярный механизм действия алкоголя на клетки мозга
Эффект от употребления алкогольных напитков известен практически каждому, однако молекулярный механизм действия этанола на мозг долгое время был совершенно не изучен - не было понятно даже, является ли действие специфическим (то есть, основанным на механизме лиганд-рецептор) или неселективным (когда этанол просто нарушает физико-химическое равновесие внутренней среды организма).
Несмотря на крайне простое строение молекулы этилового спирта - «активного вещества» этих напитков - молекулярный механизм действия алкоголя на нервную систему долгое время был совершенно неизвестен; понятно было только, что он как-то нарушает взаимодействие между нервными клетками в мозгу. Мнения колебались между двумя крайними случаями: от варианта неспецифического взаимодействия - когда этанол сдвигает физико-химический и биохимический баланс организма (гомеостаз) и влияет, в частности, на свойства клеточных мембран, - до механизма лиганд-рецепторного взаимодействия. (Последний случай подразумевает существование особого рецептора спирта, который был неизвестен, и существование которого представлялось сомнительным с учётом малого размера молекулы этанола.)
Сравнительно недавно стали появляться
биохимические данные, говорящие о чувствительности многих ионных каналов,
присутствующих в мембранах нервных клеток (например, управляемых лигандами: γ-аминомасляной
кислотой, глицином, ацетилхолином, серотонином и другими) к действию этанола и
других низкомолекулярных спиртов. Ранние гипотезы объясняли этот эффект
модификацией вязкости и текучести липидных мембран клеток, но позднее учёные
стали склоняться к идее специфических сайтов связывания молекул спиртов в этих
каналах.
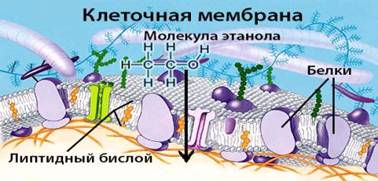
Рис. 2 Строение клеточной мембраны
Исследователи из института Солка <#"786117.files/image003.jpg">
Рис. 3. Консервативные
спирт-связывающие сайты в структурах K+-каналов входящего
выпрямления IRK1 и GIRK2 (А)
Структура цитоплазматического домена канала IRK1 в комплексе с молекулой двухатомного спирта 2-метил-2,4-пентандиола (МПД). Сайт связывания, образованный контактом двух смежных субъединиц, состоит из трёх структурных элементов: N-концевого участка, βL-βM «шпильки» одной субъединицы и βD-βE «шпильки» другой субъединицы. (На врезке схематично изображена структура одной субъединицы IRK-канала, включая поровую петлю, два трансмембранных сегмента и цитоплазматический домен, изображённый на этой картинке [показан пунктиром]). (Б, В). Детальное представление спирт-связывающих сайтов в комплексе с МПД в структуре канала IRK1 (Б) и в полученной по аналогии модели комплекса с GIRK2 (В). (Предполагаемое положение МПД во втором случае [показано пунктиром] получено путём совмещения структур цитоплазматических доменов IRK1-МПД и GIRK2.) (Г). Выравнивание аминокислотных последовательностей IRK1 и GIRK2 в области трёх структурных доменов, образующих сайт связывания спиртов (остатки, непосредственно формирующий сайт, обведены в рамку). (Д). Вольт-амперная характеристика канала GIRK2: базальный ток и активированный ток в присутствии 100 мМ МПД, составляющий ~246% от базального. Для сравнения приведён ток канала, ингибированного 1 мМ ионами Ba2+ (чёрным).
Данные об активации каналов GIRK спиртами с числом атомов углерода до четырёх (метанолом, этанолом, 1-пропанолом и 1-бутанолом) и блокировании более длинными спиртами говорят об ограниченном объёме сайта связывания; эти же сведения легли в основу проведённого эксперимента по характеристике этого сайта с помощью точечного мутагенеза.
Что интересно, схожие по форме и объёму активные сайты обнаружены и в других белках, связывающих спирты, - в частности, в алкогольдегидрогеназе, расщепляющей этанол в клетках печени, и в белке дрозофилы LUSH, ответственном за рецепцию алкоголя в окружающей среде.
Активация каналов GIRK приводит к
общему снижению активности нейронов, что согласуется с депрессорным влиянием
алкоголя на центральную нервную систему. Исследователи считают, что это
свойство может быть использовано при лечении эпилепсии, характеризующейся
эпизодическими аномальными всплесками электрической активности мозга, внешне
проявляющимися в виде печально известных припадков. «Если бы удалось
разработать новое соединение, конкурирующее с алкоголем за связывание в
активных сайтах GIRK-каналов, этим можно было бы подавлять вспышки ненормальной
активности нейронов и, возможно, получить новое лекарство от эпилепсии», -
говорит Пол Слезингер, руководитель исследования.
2.1.2 Нейрофизиология формирования алкоголизма
Нервная система человека обладает важным свойством, позволяющим легко формировать и закреплять привычки независимо от того, полезны они или вредны для здоровья. С физиологической точки зрения, приобретение привычки связано с образованием в мозговых структурах устойчивых нервных связей, обусловливающих готовность к действию.
Ткани мозга человека обладают электрической активностью, регистрируемой электроэнцефалографом. Эта активность является общим проявлением электрической активности отдельных нервных клеток. Она связана с существованием электрического потенциала мембраны клеток. Этот потенциал обусловлен различием концентрации ионов по обе стороны мембраны. Поляризация электрического потенциала клеток организма практически не меняется за исключением нервных и мышечных, в которых происходит деполяризация. Именно это свойство делает их способными передавать и принимать сигналы, т.е. возбуждаться.
Нейроны, выполняющие важнейшие функции в организме (получение, проведение и передача информации другим клеткам в виде импульсов), имеет особое строение. Нервная система состоит из электрически изолированных нейронов, но действует она как единый проводящий путь. Такая ее способность обеспечивается особым контактом между передающими нейронами и клетками-мишенями, получающими информацию (синапс). Строение синапса изображено на рис. 4.
Нейрофизиология формирования привычки
определяется образованием синаптической связи с повышенной готовностью синтеза
медиатора и чувствительностью к нему рецептора. Таким образом, в основе
передачи нервного импульса через синапс лежит единство двух противоположных
химических процессов - активация нейромедиатора и подавление его ферментом. Так
обеспечивается высокая динамичность прохождения сигнала и готовность получения
нового в информационной сети нервной системы.
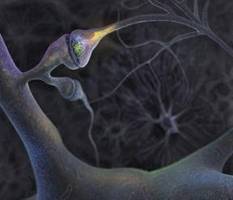
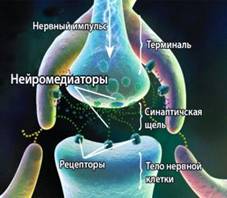
Рис. 4. Внешний вид и строение
синапса
В работе нервной системы участвует целый спектр разнообразных по химической природе эндогенных нейромедиаторов и рецепторов, отличающихся по характеру действия. Существуют семейства тормозящих и возбуждающих медиаторов.
Как свидетельствуют современные научные факты, нейрофизиологические механизмы развития зависимости базируются в стволовых и лимбических структурах мозга, в тех его областях, где располагается так называемая система подкрепления (см. рис. 5). Эта система участвует в обеспечении регуляции эмоционального состояния, настроения, мотивационной сферы, психофизического тонуса, поведения человека в целом, его адаптации к окружающей среде. В свое время было показано, что если при вживлении в данные «зоны» микроэлектродов животное получает возможность произвольно раздражать их электрическим током, то оно делает это безостановочно в течение длительного времени - вплоть до полного истощения.
Несомненно, что психоактивные
вещества, обладающие наркогенным потенциалом, т.е. способные привести к
развитию зависимости, также воздействуют химическим путем на указанную систему
подкрепления, активируя ее и влияя на метаболизм нейромедиаторов.
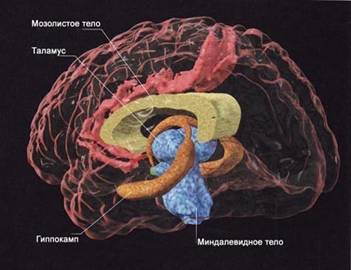
Рис. 5. Строение лимбической системы мозга
Результаты многочисленных исследований позволяют сделать заключение, что именно влияние алкоголя и наркотиков на нейрохимические процессы мозга является основой развития синдрома зависимости. При этом следует отметить, что массивное воздействие этих препаратов приводит к дисфункции почти всех нейрохимических систем мозга, однако далеко не все из этих нарушений имеют связь с развитием синдрома зависимости.
К настоящему времени накоплен убедительный материал о множественности действия этанола на функции нейронов: изменяются генерация потенциалов нервной клетки, ионные мембранные токи, рецепторный аффинитет, высвобождение нейромодуляторов и трансмиттеров. При хроническом алкоголизме существенные, порой необратимые сдвиги происходят в метаболизме нервной клетки. Так, дефицит витамина В1 (тиамина) - кофактора нескольких ферментов, затрудняет использование глюкозы нервной тканью, что является одной из причин серьезного повреждения мозга - алкогольной энцефалопатии.
Развитие устойчивости и физической зависимости - это, по-существу, процесс адаптации, которым организм отвечает на воздействие этанола как на длительное введение любого другого эндогенного или экзогенного агента. Адаптивные изменения затрагивают структуру клеточных мембран (повышается их ригидность), функции ионных каналов, белков, вторичных внутриклеточных мессенджеров, медиаторную передачу. Наиболее подробно и обстоятельно изучено влияние этанола на нейромедиаторные системы - катехоламин-, серотонин- и ГАМК-ергическую.
Анализ обнаруженных эффектов этанола на эти системы позволил выдвинуть нейромедиаторную гипотезу формирования алкогольной зависимости. Однократный прием этанола стимулирует секрецию норадреналина и дофамина из синаптических окончаний, тормозит их обратный захват и снижает активность расщепляющего катехоламины энзима (КОМТ). Согласно гипотезе, возрастание уровня катехоламинов в синаптической щели вызывает субъективное ощущение эйфории, снимает эмоциональную напряженность и является фактором внутреннего вознаграждения. Спустя некоторое время увеличивается активность α-адренорецепторов и высвобождение катехоламинов уменьшается, усиливается их разрушение ферментом пресинаптических окончаний - моноаминооксидазой (МАО). Образовавшийся таким образом дефицит катехоламинов ощущается как дискомфорт и побуждает к повторяющемуся приему этанола.
Постоянная недостаточность норадренергической системы вследствие повышения активности разрушающих катехоламины ферментов, увеличения обратного захвата и снижения чувствительности адренорецепторов характеризует состояние физической зависимости от этанола. При хроническом алкоголизме в первые сутки после прекращения приема алкоголя происходит чрезмерный, ненормированный вследствие нарушения обратной связи выброс катехоламинов, особенно дофамина. Концентрация дофамина в крови и спинномозговой жидкости возрастает в десятки раз и развивается клиническая картина абстинентного синдрома. Предполагается, что наблюдаемый в это время тремор и неадекватные движения, похожие на паркинсонизм, являются следствием нарушения дофаминергической передачи в базальных ядрах головного мозга.
Таким образом, норадренергическая система играет особую роль на всех этапах алкоголизма, обусловливая положительное эмоциональное состояние и эйфорию. Дофаминергическая система, по-видимому, не участвует в формировании алкогольной мотивации, но выступает на первый план при абстинентном синдроме.
Близкие изменения при алкоголизме претерпевает и обмен в мозге другого биогенного амина - серотонина. У предрасположенных к потреблению этанола крыс концентрация серотонина в ряде структур мозга снижена. При введении крысам ингибитора обратного захвата серотонина-флуоксетина, повышающего содержание серотонина, потребление этанола уменьшается. При синдроме отнятия активность серотонинергической системы возрастает и может быть причиной наблюдаемых в этот период галлюцинаций.
В процессе формирования зависимости от алкоголя обмен ацетилхолина в нервной ткани, вероятно, не меняется, однако при глубоких поражениях ЦНС - алкогольной амнезии - отмечают редукцию холинергических нейронов коры мозга.
В последнее время все большее внимание исследователей привлекает участие в реализации эффектов этанола ГАМК-рецепторного ионофорного комплекса. Оказалось, что этанол непосредственно усиливает ток иона хлора через мембранный канал, т. е. увеличивает ГАМК-ергическую тормозную передачу. Антагонисты ГАМКд-рецептора (пикротоксин, бикукулин) снимают седативное действие этанола на поведение и координацию движений. Этанол действует и на бензодиазепиновый рецептор этого комплекса, повышая сродство рецептора к диазепаму. В целом эти данные объясняют «барбитуратные» (седативные и анксиолитические) свойства этанола и повышение уровня алкогольной мотивации в стрессорных условиях. При поступлении одновременно с алкоголем в организм бензодиазепинов потребление этанола снижается.
Электрохимическими и нейрохимическими методами установлено, что этанол изменяет также катионные мембранные токи. Изучение проводимости потенциалзависимых Са++- каналов в присутствии этанола дало ценный материал для разработки теоретического аспекта проблемы алкоголизации и для практической медицины. Однократная обработка этанолом ингибирует вызванный деполяризацией вход ионов кальция в нейроны. Постепенно при адаптации к алкоголю поступление Са++ внутрь нейронов увеличивается, в результате чего при абстинентном синдроме потенциалзависимые Са++-каналы резко активируются и наблюдается возбуждение ЦНС, купируемое применением антагонистов кальциевых каналов. Немаловажно также то, что Са++ участвует не только в электрических процессах, но и являясь внутриклеточным мессенджером, регулирует фосфорилирование белков, секрецию гормонов и трансмиттеров.
Сведения об участии пептидергических систем организма в развитии алкоголизма противоречивы. Несмотря на то, что показано вовлечение целого ряда пептидов в регуляцию влечения к этанолу (ангиотензина II, брадикинина, тиролиберина и др.), более или менее определенные корреляции найдены лишь между уровнем потребления и содержанием опиоидных пептидов, а также аффинитетом опиатных рецепторов. Активация опиоидергической системы, как полагают, составляет существенное звено механизма внутреннего подкрепления при приеме алкоголя. У животных, предрасположенных к алкоголизму, отмечаются низкая концентрация метионин-энкефалина в некоторых отделах мозга и измененная реактивность эндорфинергической системы гипофиза. Прием этанола повышает содержание мет-энкефалина в мозге и эндорфина в спинномозговой жидкости и крови. Аналогичный эффект алкоголь оказывает на другой антистрессорный пептид - пептид дельта-сна, уровень которого в коре мозга животных с влечением к этанолу также снижен. Введение внутрь желудочков мозга метэнкефалина, лейэнкефалина, пептида дельта-сна предотвращает формирование алкогольной мотивации. По-видимому, недостаточность пептидных стресспротективных систем мозга усиливает тягу к этанолу, который в свою очередь мобилизует защитные противострессорные механизмы.