Материал: Донсков С.И., Мороков В.А. Группы крови человека. Руководство по иммуносерологии
Глава 5.
Система Kell
Антигены системы Kell (Келл) (табл. 5.1) разделяют на 4 группы.
1-я группа – 11 антигенов, объединенных в 5 подгрупп: K и k; Kp a, Kp b и
Kp c; Js a и Js b; K11 (Cote) и K17 (Wk a); K14 (San) и K24 (Cls). Аллельная взаи-
мосвязь генов внутри подгрупп четко установлена.
2-я группа – 7 часто встречающихся антигенов: K12 (Boc), K13, L18, K19 (Sub), K22, TOU и RAZ, не имеющих антитетичных партнеров.
3-я группа – 3 редко встречающихся антигена: Ul a, K23 и VLAN, молекулярная основа которых расшифрована и представляет собой простые аминокислотные замены.
4-я группа – 3 часто встречающихся антигена: Ku, k-like и Km. Молекулярная основа этих антигенов не ясна. Для того чтобы установить аминокислотную последовательность, обусловливающую их специфичность, необходимо сравнить два Kell-полипептида: содержащий и не содержащий указанные антигены. Однако лица, лишенные этих антигенов, практически не встречаются.
Kell-фенотип зависит от двух различных генных локусов. Синтез антигенного вещества Kell осуществляет локус KEL, который расположен на хромосоме 7. Экспрессию антигенов Kell контролирует другой локус – XK, находящийся на Х-хромосоме. Ген XK продуцирует Kx-протеин, который является подложкой и одновременно составной частью Kell-гликопротеина (рис. 5.1).
Локус KEL в отличие от локусов D и CE системы резус более компактен. Кроссинговер в нем не наблюдается, генные конверсии редки. Один гаплотип KEL продуцирует несколько часто встречающихся (общих для всех людей) Kellантигенов, но не более одного редко встречающегося.
Антигены Kell расположены на Kell-гликопротеине (трансмембранном гликопротеине CD238), имеющем мол. массу 93 кДа. Основная часть Kell-антигенов размещается в экстрацеллюлярной части Kell-гликопротеина (см. рис. 5.1), что, по-видимому, и делает его столь иммуногенным.
Гликопротеины Kell относятся к цинксодержащим металлопротеинам. Они обладают свойствами эндопептидаз и имеют некоторое сходство с так называемым общим антигеном острого лимфобластного лейкоза.
Молекулярный субстрат большинства Kell-антигенов известен. Их серологический полиморфизм обусловлен простыми аминокислотными заменами в Kellгликопротеине (см. табл. 5.1).
Система Kell тесно связана с двумя другими независимыми системами – Kx
341
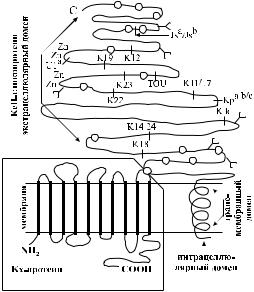
Рис. 5.1. АрхитектоникаKell-гликопротеинаиKx-протеина(гипотетическаясхемапоHoи соавт.[198]идр.авторам).
О – цистеиновые остатки,
 – цистеиновый остаток – место соединения Kell-гликопротеина и Kx-протеина,
– цистеиновый остаток – место соединения Kell-гликопротеина и Kx-протеина,
 – участки присоединения N-гликанов (гликозилирования),
– участки присоединения N-гликанов (гликозилирования),
| – позиции аминокислотных замен, определяющие специфичность антигенов Kell, Zn – цинксвязывающие участки.
и Gerbich. От последних зависит степень выраженности антигенов Kell на поверхности клетки.
Описаны редкие фенотипы, в которых большинство Kell-антигенов слабо выражены или отсутствуют (фенотипы McLeod, Kmod, транзиторный K + или K −, Ko). Исключение представляет антиген Ku, который присутствует на всех эритроцитах, в том числе на Kell-дефицитных и Ko.
Отсутствие Kell-антигенов и ослабленная их экспрессия могут быть обусловлены несколькими причинами: 1 – гомозиготностью по молчащему гену KEL-локуса; 2 – эпистатическим подавлением KEL-локуса – эпистазией генов Gerbich; 3 – блокадой антигенных участков Kell-гликопротеина аутоантителами (транзиторный K −) или бактериальной модификацией (транзиторный K + ); 4 – нарушением синтеза Kx-протеина и другими, не вполне ясными причинами. Дефицитные Kell-фенотипы передаются по наследству (Kmod, McLeod), но, по-видимому, могут формироваться и как следствие соматических мутаций.
Наибольшей иммуногенностью обладает фактор K (синонимы: Kell, KEL1, K1). Его определение регламентировано нормативными документами [54, 55] и является функциональной обязанностью иммуносерологов службы крови.
Антигены Kell представляют также интерес для антропологов [13] и судебных медиков [3].
342
|
|
|
|
|
|
Таблица 5.1 |
|
|
Антигены системы KEL |
|
|
||
|
|
|
|
|
|
|
Обозначение |
|
Частота среди |
|
Молекулярная |
Год открытия |
|
авторское |
традиционное |
ISBT |
европеоидов, % |
|
основа |
[источник] |
|
|
|
|
|
|
|
Kelleher (Kell) |
K |
KEL1 |
9 |
|
Met 193Thr |
1946 [133] |
|
|
|
|
|
|
|
Cellano |
k |
KEL2 |
99,8 |
|
Thr 193 Met |
1949 [249] |
|
|
|
|
|
|
|
Penney |
Kp a |
KEL3 |
2,3 |
|
Trp 281Arg / Gln |
1957 [87] |
Rautenberg |
Kp b |
KEL4 |
> 99,9 |
|
Arg 281Trp / Gln |
1958 [88] |
Peltz (Ko) |
Ku |
KEL5 |
> 99,9 |
|
Не установлена |
1961 [134] |
Sutter |
Js a |
KEL6 |
1;унегров19,5 |
|
Pro 597 Leu |
1959 [176] |
Mattews |
Js b |
KEL7 |
> 99,9 |
|
Leu 597 Pro |
1963 [183] |
|
K w |
KEL8* |
5,0 |
|
|
1965 [106] |
Claas |
KL |
KEL9* |
99,0 |
|
|
1968 [379] |
|
|
|
|
|
|
|
Karhula |
Ul a |
KEL10 |
< 0,1; |
|
Val 494 Glu |
1968 [169] |
у финнов 2,6 |
|
|||||
Cote |
|
KEL11 |
> 99,9 |
|
Val 302Ala |
1976 [185] |
|
|
|
|
|
|
|
Bockman |
Boc |
KEL12** |
> 99,9 |
|
His 548Arg |
1973 [272] |
|
|
|
|
|
|
|
Sgro |
|
KEL13** |
> 99,9 |
|
Не установлена |
1974 [262] |
|
|
|
|
|
|
|
Santini |
San |
KEL14 |
> 99,9 |
|
Arg 180 |
1973 [195] |
|
Pro / His / Cys |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Kx |
KEL15* |
> 99,9 |
|
Не установлена |
1971 [273] |
|
k-like |
KEL16 |
99,8 |
|
Не установлена |
1975 [267] |
|
|
|
|
|
|
|
Weeks |
Wk a |
KEL17 |
0,3 |
|
Ala 302Val |
1974 [356] |
Marshall |
|
KEL18** |
> 99,9 |
|
Arg 130Trh / Gln |
1975 [98] |
|
|
|
|
|
|
|
Sublett |
|
KEL19** |
> 99,9 |
|
Arg 492 Gln |
1979 [234] |
|
|
|
|
|
|
|
|
Km |
KEL20 |
> 99,9 |
|
Не установлена |
1968 [377] |
|
|
|
|
|
|
|
Levay |
Kp c |
KEL21 |
< 0,1 |
|
Gln 281Arg / Trp |
1979 [403] |
Ikar |
|
KEL22** |
> 99,9 |
|
Ala 322Val |
1982 [96] |
|
|
|
|
|
|
|
|
|
KEL23** |
< 0,1 |
|
Arg 382 Gln |
1987 [270] |
|
|
|
|
|
|
|
|
Cls |
KEL24 |
2,0 |
|
Pro 180Arg |
1985 [160] |
|
|
|
|
|
|
|
|
VLAN |
KEL25 |
< 0,1 |
|
Arg 248 Gln |
1996 [215] |
|
|
|
|
|
|
|
|
TOU |
KEL26 |
> 99,9 |
|
Arg 406 Gln |
1995 [213] |
|
|
|
|
|
|
|
|
RAZ |
KEL27 |
> 99,9 |
|
Glu 299 Lys |
1994 [148] |
|
|
|
|
|
|
|
|
VONG |
KEL28 |
|
|
|
2003 [*] |
|
|
|
|
|
|
|
|
KALT |
KEL29 |
|
|
|
2006 [*] |
|
|
|
|
|
|
|
343
|
|
|
|
|
Окончание табл. 5.1 |
|
|
|
|
|
|
|
|
|
Обозначение |
|
Частота среди |
Молекулярная |
Год открытия |
|
авторское |
|
традиционное |
ISBT |
европеоидов, % |
основа |
[источник] |
|
|
|
|
|
|
|
|
|
KTIM |
KEL30 |
|
|
2006 [*] |
|
|
|
|
|
|
|
|
|
KYO |
KEL31 |
|
|
2006 [*] |
|
|
|
|
|
|
|
|
|
KUCI |
KEL32 |
> 99,9 |
|
2007 [*] |
|
|
|
|
|
|
|
|
|
KANT |
KEL33 |
> 99,9 |
|
2007 [*] |
|
|
|
|
|
|
|
|
|
KASH |
KEL34 |
> 99,9 |
|
2007 [*] |
|
|
|
|
|
|
|
|
|
Ko (null) |
|
|
|
1957 [125] |
|
|
McLeod |
|
Очень редко |
|
1961 [86] |
|
|
|
|
|
|
|
KEL- |
|
Leach |
|
|
|
1985 [204] |
|
|
|
|
|
|
|
|
Mullins |
|
|
|
1988[308] |
|
дефицитные |
|
|
|
|
||
|
|
|
|
|
|
|
фенотипы |
|
Allen |
|
|
|
1993 [147] |
|
|
|
|
|
|
|
|
|
Kmod |
|
|
|
1995 [322] |
|
|
Транзиторный |
|
Редко |
K − стали K + |
[278] |
|
|
K +/K − |
|
K + стали K − |
[344,380,397] |
|
|
|
|
|
|||
|
|
|
|
|
|
|
* Kell-антигены, исключенные из системы Kell, ** Антигены пара-KEL,
[*] См. гл. 37 раздел «Системы Kell и Kx».
Номенклатура
До 1961 г. было принято буквенное обозначение антигенов Kell: K, k, Kр а, Kр b, Js а и т. д. В 1961 г. Allen и Rosenfield [89] предложили буквенно-цифровое обозначение: K1 (K), K2 (k), K3 (Kр а), K4 (Kp b) и т. д., которое было положено в основу международной классификации.
В соответствии с рекомендациями номенклатурного комитета Международного общества трансфузиологов – International Society of Blood Transfision (ISBT) с 1985г.дляобозначенияуказаннойсистемыпринятсимволKEL.
По классификации ISBT каждой системе антигенов эритроцитов присвоен трехзначный код, каждому антигену – трехзначный номер, например: си-
стеме АВО – 001, антигену А – 001001, В – 001002, MNSs – 002, Rh-Hr – 004, Lutheran – 005 и далее в хронологическом порядке их открытия. Системе антигенов KEL присвоен код 006. Антиген K обозначается как 006001, антиген k – 006002, антигены Пенни и Раутенберг соответственно 006003 и 006004 и т. д. В повседневной практической работе чаще используют буквенное (K, k, Kp a, Kp b) или буквенно-цифровое (K1, K2, K3, K4) обозначение (см. табл. 5.1).
Некоторые антигены после уточнения переведены в другие антигенные системы. Так, антигены K w, KL, Kx, причисленные сначала к системе KEL, из нее выведены, поскольку относятся к другим антигенным системам.
344
По правилам ISBT при перемещении антигена из одной системы в другую или исключении из номенклатуры его порядковый номер остается свободным и не используется для обозначения каких-либо других антигенов. В связи с этим в номенклатуре системы KELимеются пропуски – номера 008, 009, 015, первоначально присвоенные антигенам K w, KLи Kx соответственно.
Аргументы в пользу шестизначного обозначения ISBT, пригодного для компьютерной обработки, неоспоримы, однако традиционное обозначение более удобно и информативно для большинства иммуносерологов. Антитетическая связь между антигенами более наглядна при обозначении K и k, Kp a и Kp b, Js a и
Js b,чем K1 и K2, K3 и K4, K6 и K7.
K и k
В1945 г., вскоре после открытия резус-фактора, англичане Coombs, Mourant
иRace [132] разработали антиглобулиновую пробу, получившую название пробы Кумбса. Фактор Kell (K) был одним из первых антигенов, открытых с помощью этого метода.
Анализируя причину желтухи у новорожденного одной из родильниц (мис-
сис Kelleher), Coombs, Mourant и Race [133] обнаружили необычные анти-
тела, которые нельзя было отнести к системе резус. Сыворотка крови г-жи Kelleher реагировала с эритроцитами мужа и ребенка, а также эритроцитами примерно 7 % произвольно взятых лиц независимо от их групповой- и резуспринадлежности. Новый фактор эритроцитов получил название Kell (K) – по фамилии носительницы антител.
Позднее антитела такой же специфичности выявили Wiener, Sonn-Gordon [396] в сыворотке крови больных, перенесших гемотрансфузионные осложнения.
Внастоящее время обнаружение анти-K-антител не является редкостью. Антиген K – сильный иммуноген, и даже одна трансфузия эритроцитов, одна беременность или один аборт могут вызвать аллоиммунизацию [7, 21, 79]. По иммуногенности фактор K стоит на втором месте после D. Частота анти-Kell- антител среди аллоиммунизированных составляет более 5 %, что еще раз подчеркивает значение фактора K в трансфузиологии и необходимость типирования доноров и реципиентов по этому антигену [20, 21, 23, 24].
Спустя 3 года после открытия Kell-фактора Levin и соавт. [249] обнаружили антитела, агглютинирующие эритроциты 99,8 % лиц, и установили антитетичную связь определяемого с их помощью антигена с антигеном Kell. Второй антиген был назван Cellano (k) (в русской транскрипции Челлано) также по фамилии женщины, в крови которой были выявлены антитела. Аллельность генов K
иk подтверждена популяционными и посемейными исследованиями [250, 251, 318, 339]: K и k являются продуктом кодоминантных аллелей. Лица, не содержащие антигена K, всегда содержат k и, наоборот, лица, не имеющие k, содержат K. Оба антигена могут присутствовать на эритроцитах вместе – фенотип Kk. Отсутствие обоих антигенов (фенотип Ko) встречается редко и, как правило,
345