Материал: Донсков С.И., Мороков В.А. Группы крови человека. Руководство по иммуносерологии
Предположение о том, что полипептиды Rh могут быть связаны в мембране эритроцитов с гликозилированным компонентом (гликопротеин Rh), было впервые высказано Gahmberg [295]. Далее было установлено, что гликопротеин Rh, преципитированный анти-D-антителами вместе с полипептидом Rh, имеет мол. массу 45–70 кДа, а полипептид Rh – 30 кДа.
Когда стало ясно, что полипептиды Rh связаны в клеточной мембране с гликопротеинами Rh, появились высказывания, что одни антигены Rh могут быть экспрессированы на полипептиде, а другие – на гликопротеине. Не исключали и третий вариант: некоторые антигены Rh представляют собой комплекс, состоящий из участков полипептида и гликопротеина. Присутствуя порознь, эти участки полипептида и гликопротеина не являются иммуногенными, а когда присутствуют одновременно, их комплекс приобретает иммуногенность.
Однако в последующих работах было установлено, что белковые последовательности, определяющие специфичность антигенов Rh, расположены на полипептидах, а не на гликопротеинах.
Структура полипептидов Rh
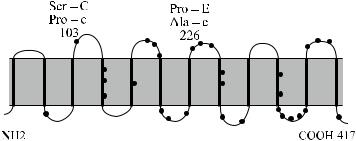
Антигенные детерминанты Rh расположены на негликозилированных нефосфорилированных полипептидах с мол. массой 30–32 кДа [129, 295, 483]. Полипептиды RhD и RhcE представляют собой цепь из 12 связанных со скелетом мембраны доменов [348], пересекающих мембрану эритроцита от эндодо экзоцеллюлярного уровня (рис. 4.5 и 4.6). Основная часть полипептида размещена в фосфолипидном бислое. На внешней стороне клетки домены соединены 6 выступающими над поверхностью мембраны петлями, на которых также могут располагаться серологически выявляемые Rh-антигены. N- и C-концевые участки полипептида погружены внутрь клетки [196]. Полипептид RhD, полученный искусственно в трансфектных клетках с помощью олигонуклеотидных праймеров и соответствующих кДНК людей Rh +, состоит из 417 аминокислот [147, 172, 590]. Из такого же числа аминокислот состоит полипептид RhcE, полученный таким же способом с использованием кДНК людей Rh − [138, 390, 418].
Рис. 4.5. Топология мембранных доменов Rh-Hr по Schenkel-Brunner [597].
176
Полипептид сЕ имеет 6 цистеиновых остатков, 5 из которых расположены в цитоплазматических петлях. Шестой цистеин (Цис-285) находится в 5-й внеклеточной петле [564]. Цистеин в позиции 311 полипептида RhcE замещен на тирозин в полипептиде RhD. Отдельные последовательности (мотивы), например Cys – His – Leu – Ile–Proвположении285–289,являютсяобщимидлявсехэпитопов:D,CcиEe.
Высокая степень гомологии между генами RHD и RHCE способствует генной конверсии, неравновесному кроссинговеру и образованию в результате этого гибридных генов, кодирующих продукцию новых антигенов Rh [597].
Rh-протеины высокогидрофобны и весьма прочно соединены с другими гидрофобными белками мембраны [336]. Обнаружена определенная связь Rh-полипептидов с гликофорином В, антигенами LW, гликопротеином, несу щим антигены Duffy, гликопротеином CD47 и так называемым Rh-ассоцииро ванным гликопротеином.
Rh-антигены устойчивы к воздействию протеолитических ферментов [295]. Антигены D и c (hr') разрушаются под воздействием N-этилмалеинимида, хлормеркурибензоната и 2-нитробензойной кислоты. Это послужило для исследователей основанием полагать, что Rh-субстанция содержит тиоловые группы
[316, 319, 600, 646].
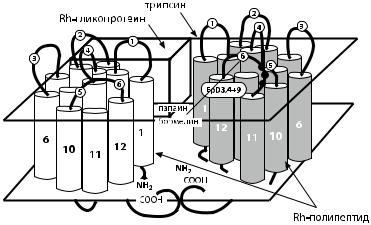
Рис. 4.6. Предполагаемая трехмерная структура доменов Rh-полипептида и Rhассоциированногогликопротеинавмембранеэритроцита(поAvent[141]).Светлыеитем- ныецилиндрыпредставляютдоменыRh-полипептидовDиСЕсN-иC-терминальными группами. На заднем плане условно представлен Rh-ассоциированный гликопротеин. Указаны участки разрывов при действии трипсина, папаина или бромелина, место расположения эпитопов D3, D4 и D9, экзо- и эндоцеллюлярные петли Rh-полипептида.
Как отмечали Dahr и соавт. [242], серологическая активность D-антигена утрачивалась под действием цистеиновых реагентов. После обработки эритроцитов дитионитробензойной кислотой (ДТНБК) антиген D инактивировался , однако активность его вновь восстанавливалась под действием дитиоэритритола. В то же время под действием глютатиона активность D-антигена не
177
восстанавливалась. Обработка эритроцитов хлормеркурифенилсульфониковой кислотой приводила практически к полной потере активности антигена D, которая не восстанавливалась дитиоэритритолом.
Йодацетамид не инактивировал D-антиген – это согласовывалось с предположением Dahr и соавт. о том, что цистеиновые остатки являются непосредственной составной частью D-антигена. Авторы пришли к выводу, что цистеиновая модификация Rh-протеина, в частности Cys 285 в 5-й внеклеточной петле, приводит к модификации антигена Rh.
Серологическая активность Rh-антигенов, как показал Green [317, 318], во многом зависела от содержания липидов в мембране эритроцитов. После вытяжки липидов из стромы n-бутанолом Rh-активность утрачивалась, а после инкубации стромы с липидным экстрактом она полностью восстанавливалась. Как указывает Schenkel-Brunner [597], липиды необходимы для оптимальной пространственной ориентации других структурных молекул в мембране эритроцита.
Обработка эритроцитов фосфолипазой, расщепляющей жирные кислоты, лектином, фосфатидилэтаноламином или фосфатидилсерином выраженно ингибировала активность антигенов c, D и e [320, 3366, 530].
На серологическую активность Rh-антигенов влияло обезвоживание мембраны эритроцитов [154, 181, 609]. Высокий уровень холестерола (соотношение холестерола и фосфолипидов 1,55) совпадал с повышенной вязкостью мембраны и большей выраженностью D-антигена. Низкий уровень холестерола (соотношение холестерола и фосфолипидов 0,55) сопровождался меньшей вязкостью мембраны и менее активным реагированием D-антигена [597].
Отсутствие Rh-полипептидов у людей с фенотипом Rhnull сопряжено с изменениями в структурной организации липидного слоя мембраны и нарушением водно-ионного транспорта в клетке.
Из ранних работ (до 1960 г.) известно, что резус-антиген термолабилен и слабеет при высушивании (П.Н. Косяков [69]). Сыворотки антирезус снижали свою активность при смешивании со стрептомицином, дериватами рибонуклеиновой кислоты, некоторыми сахарами и другими химическими веществами, из чего авторы делали предположения о возможной химической природе резус-антигена.
Наличие в эритроцитах Rh + Rh-ассоциированного гликопротеина, повидимому, вводило в заблуждение исследователей, полагавших, что антигены резус имеют полисахаридную природу.
Молекулярно-биологические исследования
Целью молекулярно-биологических исследований было ответить на вопрос: находятся ли антигенные детерминанты D, С / с и Е / е на разных белковых структурах или же они присутствуют на одном и том же полипептиде? Другой целью этих исследований было установить молекулярную структуру вариантов Rh-антигенов и кодирующих их генов.
178
Эксперименты с использованием трансфекции фрагментов ДНК [196, 496, 618] показали, что детерминанты С / с и Е / е расположены на одном полипептидном продукте, а D – на другом. Дополнительные доказательства того, что антигены Сс и Ее могут находиться на одном и том же полипептиде, полученыAvent и соавт. [145] в исследованиях с сыворотками анти-С, анти-с, анти-Е и анти-е, которые, как выяснилось, реагируют с разными участками одного и того же полипептида Rh.
Антигенные детерминанты Rh кодируются двумя похожими по структуре генами (Colin и соавт. [233], Arce и соавт. [138]). Один из них (RHD) определяет наличие трансмембранного белка, придающего эритроцитам D-актив ность. У лиц Rh + этот ген представлен одной или двумя копиями. Как уже упоминалось, у большинства лиц Rh − ген, кодирующий субстрат D, отсутствует. Находящийся в прилежащем локусе ген RHCE определяет экспрессию антигенов С, с, Е и е.
Гены RH в каждом гаплотипе с большой точностью контролируют количество вырабатываемого антигенного вещества. У разных членов одной и той же семьи гаплотип RH кодировал продукцию одинакового количества каждого присутствующего в фенотипе антигена (Rosenfield, Kochwa [393, 576]).
Принцип идентификации генов, ответственных за продукцию Rh-антигенов, сводится к следующему:
1)выделение несущего Rh-активность белка с помощью преципитации специфическими антителами;
2)расшифровка аминокислотной последовательности иммунодоминантного белка, приготовление комплементарных молекулярных зондов и конструкций для идентификации кодирующей ДНК (кДНК);
3)вживление (трансфекция) кДНК в клетки или плазмиды, не производящие аналогичного иммунодоминантного продукта;
4)идентификация геномного продукта трансфектных клеток или плазмид;
5)идентификация геномной ДНК (гДНК), представляющей собой собственно ген с кодовой записью иммунодоминантного продукта.
Клонирование Rh-полипептидов
В 1990-х годах 2 лаборатории независимо друг от друга получили совершенно одинаковые клоны кДНК полипептидов RhcЕ (Cherif-Zahar и соавт. [208], Avent и соавт. [148]).
Выделение кДНК, соответствующей полипептиду D, было осуществлено независимо 3 исследовательскими группами (Le Van Kim и соавт. [418],Arce и соавт. [138], Kajii и соавт. [390]). Последовательность аминокислот в белках, полученных при помощи кДНК RhD, отличалась от аминокислотной последовательности полипептида RhсЕ по 34–36 аминокислотным остаткам из 417 последовательностей (рис. 4.7).
179
D |
|
|
|
|
|
|
MSSKYPRSVR |
RCLPLWALTL |
|
cE |
|
|
|
|
|
50 |
- - - - - - - - - - |
- - - - - - - - - - |
|
|
|
|
|
|
|
|
|
|
|
D |
EAALILLFYF |
FTHYDASLED |
QKGLVASYQV |
GQDLTVMAAI |
GLGFLTSSFR |
||||
cE |
- - - - - - |
- - - - |
- - - - - - - - - - |
- - - - - - - - - - |
- - - - - - - - - L |
- - - - |
- - - N- - |
||
|
|
|
|
|
|
100 |
|
|
|
D |
RHSWSSVAFN |
LFMLALGVQW |
AILLDGFLSQ |
FPSGKVVITL |
FSIRLATMSA |
||||
cE |
- - - - - - |
- - - - |
- - - - - - - - - - |
- - - - - - - - - - |
- - P- - - - - - - |
- - - - - - - - - - |
|||
|
|
|
|
|
|
150 |
|
|
|
D |
LSVLISVDAV |
LGKVNLAQLV |
VMVLVEVTAL |
GNLRMVISNI |
FNTDYHMNMM |
||||
cE |
M - - - - - |
AG- - |
- - - - - - - - - - |
- - - - - - - - - - |
- T - - - - - - - - |
- - - - - - - - LR |
|||
|
|
|
|
|
|
200 |
|
|
|
D |
HIYVFAAYFG |
LSVAWCLPKP |
LPEGTEDKDQ |
TATIPSLSAM |
LGALFLWIFW |
||||
cE |
-F - - - - |
- - - - |
-T - - - - |
- - - - |
- - K - - - - |
N - - |
R - - - - - - - - - |
- - - - - - - M - - |
|
|
|
|
|
|
|
250 |
|
|
|
D |
PSFNSALLRS |
PIERKNAVFN |
TYYALVVSVV |
TAISGSSLAH |
PQGKISKTYV |
||||
cE |
- - V- - P |
- - - - |
- -Q- - - - |
M - - |
- - - - - A - |
-300- - |
- - - - - - - - - - |
- - R |
- - -M - - - |
|
|
|
|
|
|
|
|
|
|
D |
HSAVLAGGVA |
VGTSCHLIPS |
PWLAMVLGLV |
AGLISVGGAK |
YLPGCCNRVL |
||||
cE |
- - - - - - |
- - - - |
- - - - - - - - - - |
- - - - - - - - - - |
- - - - -I - - - - |
C - - |
V - - - - - - |
||
|
|
|
|
|
|
350 |
|
|
|
D |
GIPRSSIMGY |
NFSLLGLLGE |
IIYIVLLVLD |
TVGAGNGMIG |
FQVLLSIGEL |
||||
cE |
- -R - I -V- HS |
I - - - - - |
- - - - |
-T - - - - - |
- - R |
- - WN - - - - - - |
- - - - - - - - - - |
||
|
|
|
|
|
|
400 |
|
|
417 |
D |
SLAIVIALTS |
GLLTGLLLNL |
KIWKAPHEAK |
YFDDQVFWKF |
PHLAVGF |
||||
cE |
- - - - - - |
- - - - |
- - - - - - - - - - |
- - - - - - - V - - |
- - - - - - - - - - |
- - - - - - - |
|||
Рис. 4.7. Аминокислотная последовательность транскриптов гена D и cE по
Mouro и соавт. [496] и Schenkel-Brunner [597]. Пунктир означает одинаковую аминокислотную последовательность сравниваемых полипептидов.
Полипептиды D и сЕ имеют высокую степень гомологии: клон полипептида D, выделенныйArce и соавт. [138], был на 96 % идентичен клону полипептида сЕ по последовательности нуклеотидов кДНК и на 92 % идентичен по структуре белка.
Le Van Kim и соавт. [418] показали, что полученные ими клоны кДНК являются продуктом гена D, поскольку фрагменты геномной ДНК, с которыми совпадали эти клоны, были получены от людей с фенотипом D +.
В то же время Kajii и соавт. [390] установили, что полученные ими клоны не были продуктом гена D, поскольку кДНК была идентифицирована у человека с фенотипом D −C +c + E +e + [390, 665, 666].
Данные о том, что у некоторых людей фенотип D − обусловлен делецией гена D, были получены Colin и соавт. [233] в экспериментах с использованием метода Саузерн-блот.
Клонирование Rh-гликопротеинов
Moore и Green [481] обнаружили, что в процессе иммунной преципитации полипептидов D и сЕ специфическими антителами одновременно с по липептидами преципитируются N-гликозилированные белки, получившие название RhAG (Rh-ассоциированные гликопротеины), или Rh-гликопротеины . Компоненты этих белков, сопровождающие полипептиды D и еС, имели несколько отличающуюся мол. массу, но одинаковую N-концевую аминокислотную последовательность [147].
180