Материал: Донсков С.И., Мороков В.А. Группы крови человека. Руководство по иммуносерологии
Рис.4.1.Генетическиеконцепциисистемырезус. 1–теорияодногогенаA.Wiener(1943),
2–теориятрехгеновR.FisherиR.Race1946), 3–теориядвухгеновP.Tippett(1986).
Согласно представлениям Винера, разнообразие факторов резус и их сочетаний в наблюдаемых фенотипах людей обусловлено одним геном, который встречается в виде8аллеломорфныхвариантов:R1,R2,Rо,RZ,r,r',r" (см.табл.4.4).
Обнаружение новых факторов (E, e, C W и др.), легко укладывались в концепцию Винера. Так, после открытия антигена hr' (c) и hr" (e) он дополнил первоначальный постулат, указав, что ген Rо может кодировать помимо Rhо еще и факторы hr' и hr" (фенотип Rоr, или cDe). Открытие антигена C W Винер трактовал как обнаружение нового аллеля R1w, а открытие антигена hr S, присутствующего у большинства людей hr" (e), – как новый аллель R^hо.
Однако по мере открытия новых антигенов и их разновидностей обозначения Винера стали затруднительными.
К середине 50-х годов иммуносерологи располагали уже не двумя, а пятью сыворотками антирезус, дифференцирующими соответствующие антигены резус: D, C, E, c и е. С помощью этих сывороток американской школой исследователей во главесВинеромианглийскойшколойвоглавесФишеромиРейсомбылиустановлены8гаплотипов,кодирующихразличныесочетанияантигенов(см.табл.4.4).
В отличие от Винера, утверждавшего, что ген резус неделим, Фишер и Рейс полагали, что существуют 3 сцепленных локуса (теория трех генов), которые наследуются одновременно. Согласно их представлению, в этих локусах на хромосоме в линейном порядке расположены гены D, C и E, кодирующие антигены D, C и E (см. рис. 4.1). Доссе [50] добавил четвертую пару аллельных генов Ff. В этих же локусах располагаются соответствующие им 3 аллельных гена: d, c и е, кодирующие антитетичные антигены d, c и е. В каждом локусе может присутствовать один ген: D или d, C или c, E или e. Таким образом, каждый индивид получает с хромосомой матери и отца от 3 до 5 антигенных признаков, определяющих его резус-фенотип.
161
Фишер сформулировал понятие об антитетичных антигенах и предсказал существование антигенов Е, е и d. Предвидение двух первых вскоре (в 1943 г.) блестяще подтвердили Wiener, Sonn [713], открыв антиген Е, и Mourant [494] (в 1945 г.) – е. Третий гипотетический антиген (d) так и не был обнаружен.
Антитела анти-d были описаны Hill и Haberman в 1948 г. [349], Matthes в 1950 г. [467], однако эти находки не подтвердились. По-видимому, указанные авторы исследовали неизвестные в то время антитела анти-се, реагирующие с эритроцитами cde, но не реагирующие с эритроцитами CDe, cDE, что создавало видимость анти-d-специфичности. Как сегодня известно (см. далее), у лиц Rh − в локусе, симметричном D, генетический материал в виде аллеля d не найден. Отсутствие гена d объясняет, почему многочисленные целенаправленные поиски антигена d не увенчались успехом.
Теория трех генов позволила установить последовательность генов в генном локусе RH, а также объяснить происхождение редких фенотипов резус.
В соответствии с этой теорией локусы RH располагаются в последовательности Dd – Cc – Ee (см. рис. 4.1). По мнению Фишера и Рейса, разрыв генного комплекса происходит на дистанции С – E чаще, чем на дистанции D – C, из чего следует, что гены D и E наиболее удалены друг от друга. Доказательством такого расположения генов явилась находка фенотипа −D − (Race и соавт. [552], Read и соавт. [556]), для которого характерно отсутствие антигенов Сс и Ее. Если бы ген D располагался между генами C и E, Cc – Dd – Ee, то делеция локуса Cс и Eе в варианте −D − должна была так или иначе сказаться на гене D. Однако, напротив, антиген D в фенотипе −D − серологически более выражен, чем в фенотипе CDe и cDE, что связано, по-видимому, с отсутствием конкурентного влияния генов С и Е на D вследствие их делеции.
Согласно теории трех генов, редкие генные комбинации (dCe, dcE, DCE, dCE) возникают в результате кроссинговера частых генных комбинаций: DCe, DcE, dce. На рис. 4.2 приведены варианты кроссинговера. Если кроссинговер происходит между хромосомами, несущими гaплотипы DСe и dce, образуется редкий гаплотип dCe; если между хромосомами, несущими гаплотипы DcE и dce, – редкий гаплотип dcE; если между DСe и DcE – DСE. При повторном кроссинговере DCE и dce образуется еще более редкий гаплотип – dCE. Как видно из рис. 4.2, гаплотип Dce образуется при каждом из перечисленных вариантов кроссинговера. Сумма частоты (2,49 %) редко встречающихся в европеоидных популяциях фенотипов (dCe, dcE, DCE, dCE) и частота фенотипа Dce (2,6–3 %) примерно совпадали, что подтверждало правильность теории в целом.
Относительная редкость кроссинговера (≈ 3 %), по мнению Фишера и Рейса, свидетельствует о том, что гены DCE близко расположены друг к другу и кроссинговер между ними происходит скорее как исключение, чем как правило.
Большинство исследователей нашли CDE-терминологию более удобной для повседневного использования, чем Rh-Hr, и она получила статус прикладной.
162
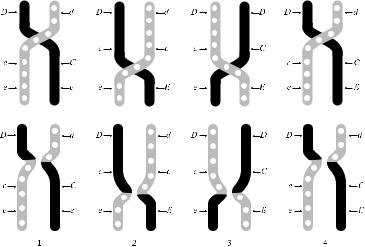
Рис. 4.2. Варианты кроссинговера по Фишеру и Рейсу.
1 и 4 – перекрест в участке D – C, 2 и 3 – перекрест в участке С – Е. В результате кроссинговера возникают следующие гаплотипы: 1 – dCe и Dce, 2 – dcE и Dce, 3 – DCE и Dce, 4 – dCE и Dce.
Благодаря своей простоте теория трех генов Фишера – Рейса завоевала всеобщее признание и несколько потеснила концепцию Винера, которая отнюдь не утратила своего значения и практического применения до настоящего времени.
Спустя годы концепцию трех тесно связанных локусов модифицировали в однолокусную, которая предполагает 3 сублокуса в одном генном комплексе.
Открытие Sanger и соавт. [596] антигенов f (ce) и rhi (Ce) выходило за рамки концепции трех генов, постулирующей принцип «один ген – один антиген». Следовало признать, что система Rh включает не 3, а 4 (C, D, E + ce), затем 5 (C, D, E, ce + Ce) локусов. Перекрестно реагирующие сыворотки и необычные Rh-фенотипы также не вписывались в теорию трех генов и кроссинговера по Фишеру. Номенклатура Фишера – Рейса стала затруднительной для обозначения большого числа новых Rh-антигенов и ей на помощь пришла цифровая номенклатура, предложенная Розенфельдом.
Свои концепции Винер, Фишер и Рейс основывали исключительно на результатах серологических исследований (популяционные и семейные). Концепция Типпетт (теория двух генов) также основана на данных серологических исследований, однако более широкого масштаба. К началу 90-х годов арсенал сывороток, которым располагали иммуносерологи, составил более 40 наименований, включая анти-се, анти-Се, анти-сЕ и анти-СЕ. Накопились многочисленные данные о качественном и количественном разнообразии фенотипов Rh, в том числе данные о биохимической природе Rh-антигенов.
Moore и соавт. [482] в 1982 г. и Ridgwell и соавт. [564] в 1983 г. независи-
мо друг от друга нашли, что Rh-антигены располагаются на двух протеинах
163
мембраны эритроцитов: один белок несет на себе антиген D, другой – C и E. Имеющиеся сведения послужили толчком к формированию новой, современной молекулярно-генетической модели системы Rh, предложенной Типпетт.
Согласно концепции Типпетт [654, 656], система Rh контролируется двумя тесно сцепленными структурными генами, один из которых кодирует D-антиген, другой – антигены С, с, Е и е. Первый ген включает 2 аллеля: D, получивший название RHD, и не-D (отсутствие кодирующего субстрата); второй ген представлен четырьмя аллелями: се, Се, сЕ и СЕ. Продукты второго гена, получившего название RHCE, идентифицируют с помощью 5 специфических сывороток: анти-D, анти-се, анти-Се, анти-сЕ и анти-СЕ (табл. 4.7). По мнению Типпетт, отдельные эпитопы полипептида СсЕе (С, с, Е и е) более иммуногенны, чем их комплексы (се, Се, сЕ и СЕ), в связи с чем антитела анти-с, анти-С, анти-Е и анти-е встречаются чаще, чем комплексные антитела анти-се, анти-Се, анти-сЕ, анти-СЕ.
Данные, представленные Типпетт (см. табл. 4.7), с такой же убедительностью свидетельствуют о существовании четырех аллелей гена СЕ, как и данные, полученные в свое время Wiener, Fisher и Race с помощью пяти специфических сывороток: анти-D, анти-с, анти-С, анти-Е и анти-е. Вопрос о том, какая концепция ближе к действительности, сегодня решается в пользу взглядов Типпетт, подтверждающихся данными молекулярно-биологических исследований.
Необычные, в том числе редкие, фенотипы резус, как полагает Типпетт, возникают в результате мутаций, делеций и транслокаций генетического вещества. На это указывают фенотипы с ослабленными антигенами (С)(е) или (с)(е) (табл. 4.8), которые ассоциированы с редкими антигенами – Rh32, Rh35, Rh36, Rh48. Мутации и другие воздействия на генный локус RH нарушают продукцию нормального антигена, создают новые необычные формы антигенов.
Таблица 4.7
Реакции генного продукта основных 8 гаплотипов*
Генный |
|
Реакция с сыворотками |
|
||
продукт |
анти-D |
анти-ce |
анти-Ce |
анти-cE |
анти-CE |
|
|
|
|
|
|
Dce |
+ |
+ |
– |
– |
– |
DCe |
+ |
– |
+ |
– |
– |
DcE |
+ |
– |
– |
+ |
– |
DCE |
+ |
– |
– |
– |
+ |
dce |
+ |
+ |
– |
– |
– |
dCe |
– |
– |
+ |
– |
– |
dcE |
– |
– |
– |
+ |
– |
dCE |
– |
– |
– |
– |
+ |
* По Tippett [654].
164
Таблица 4.8
Антигены, продуцируемые редкими аллелями RHCE-локуса при его повреждении*
Повреждение |
Продуцируемые |
СЕ протеина |
антигены |
(С)(е) |
Rh9 C x |
(С)(е) |
Rh32 |
(С)(е) |
Rh35 |
(С)(е) |
Rh48 (JAL) |
(с)(е) |
Rh36 (Be a) |
* По Tippett [654]. |
|
Задолго до Tippett (в 1964 г.) идею о существовании двух генов RH, структурного и операторного, высказал Lauer [410], однако его исследования не были продолжены.
Sanger и соавт. [596], исследуя природу антигена f, установили, что этот антиген вырабатывается в случае, если гены с и е расположены на одной хромосоме в положении цис. Такое же заключение было сделано ими относительно антигена Се: он вырабатывается цис-комбинацией генов С и е. Race и Sanger приблизились ксовременномупониманиютого,чтогенетическийматериал,именовавшийсяранее локусами с, е, С и Е, представляет собой один и тот же ген, имеющий аллели се, Се, сЕ и СЕ. Однако этот вывод не был ими сформулирован.
Следует обратить внимание на некоторые противоречия и сложности, привнесенныеновымпониманиемтого,чтосистемаRhкодируетсянетремяиличетырьмяпарамиаллельныхгенов,атолькодвумя:RHD иRHCE.Прежниегенетические теории объясняли все предельно просто. Так, в соответствии с Rh-Hr-концепцией фенотип R1R2, или Rhо' " hr' ", обусловлен гаплотипами R1 и R2; в соответствии с CDE-концепцией фенотип СсDEe кодируется генами С, с, D, E и e, переданными индивиду по наследству с гаплотипами CDe и cDE. С позиций двухгенной теории фенотип СсDEe объяснить сложнее. Согласно двухлокусной модели индивид СсDEe должен унаследовать ген RHD и один из аллелей гена RHCE (RHce, RHCe, RHcE илиRHCE).Влюбойизвозможныхкомбинаций(Dce,DCe,DcE,DCE)полного набора антигенов СсDEe не получается и в этом заключается противоречие.
Вряд ли можно полагать, что аллель RHCe производит антигены с и Е, а аллель RHcE – антигены С и е. Это маловероятно, поскольку нарушает основную идею двухгенной модели и, кроме того, не соответствует результатам серологических исследований. Остается признать, что фенотип СсDEe является продуктом гибридного гена Ce-D-cE. Такое объяснение более правдоподобно. Как показали результаты исследования последних лет, фенотип cde часто обусловлен делецией гена RHD. Возможность гибридизации генов RH, в силу их высокой гомологии, не вызывает сомнения и, по-видимому, явление частое.
165