Материал: Донсков С.И., Мороков В.А. Группы крови человека. Руководство по иммуносерологии
Антигены Н, А и В формируются трансферазами, которые присоединяют соот- ветствующиемоносахаридыкцепям-предшественникам.
Н-антиген представлен L-фукозой, присоединенной в позиции С-2 терминального галактозного остатка.
А- и В-антигены появляются в результате присоединения к фукозилированному галактозному остатку (Н-антигену) N-ацетил-D-галактозамина или D-галактозы в позиции С-3. Хотя структура Н-антигена представлена не только фукозой, данная группировка считается иммунодетерминантной, поскольку дефукозилирование приводит к утрате Н-серологической активности субстрата. Аналогичным образом N-ацетил-D-галактозамин и D-галактозу относят к иммунодетерминантным структурам, определяющим А- и В-серологическую актив-
ность соответственно (Schenkel-Brunner [195]).
Выделяют шесть типов АВН-активных цепей (типы 1 – 6). Цепи типа 1 присутствуют в секретах, плазме и тканях энтодермального происхождения. Цепями типа 2 представлено большинство АВН-активных олигосахаридов на эритроцитах и в тканях экто- и мезодермального происхождения. Тип 3 несет антигенные детерминанты в гликолипидах эритроцитной мембраны и в муцине у индивидов группы А (Anstee [68]). Тип 4 связан с гликолипидами и представлен в небольшом количестве на эритроцитах (Anstee [68], Daniels [87], Schenkel-Brunner [195]). Тип 6 присутствует в виде свободных олигосахаридов в грудном молоке и моче. Цепи 5-го типа в организме не встречаются, они синтезированы искусственно (Daniels [87], Schenkel-Brunner [195]).
Синтез Н-антигена происходит при участии α1,2-L-фукозилтрансферазы, которая обеспечивает перенос фукозы от гуанозин-дифосфата (ГДФ) к галактозному остатку цепи-предшественника в позиции С-2. Известны 2 типа α1,2-L- фукозилтрансферазы, синтез каждого из них контролируют высокогомологичные, однако генетически независимыме друг от друга локусы FUT1(H) и FUT2(SE). Они расположены на хромосоме 19. Продукты указанных генов (ферменты) обеспечивают фукозилирование и образование Н-активных структур в различных тканях. FUT1(H) обладает большей аффиностью к цепям типа 2, в то время как FUT2(SE) – к цепям 1-го типа. У подавляющего большинства людей антиген Н присутствует в обязательном порядке, Н-дефицитные фенотипы очень редки.
Н-антиген, образовавшийся в результате действия α1,2-L-фукозилтрансфераз, является субстратом для дальнейшего гликозилирования А- и В-специфическими трансферазами, обеспечивающими присоединение иммунодетерминантных группировок: N-ацетил-D-галактозамина и / или D-галактозы, после чего субстрат при- обретаетА-и / илиВ-антигенныесвойства.
Гены, контролирующие А- и В-трансферазы, независимы от локусов FUT1(H) и FUT2(SE), картированы на хромосоме 9, в локус АВО. Последний нередко содержит молчащие аллели О1, О2 и др., в присутствии которых синтеза А- и В-трансфераз не происходит. У лиц, гомозиготных по таким аллелям, вещество Н не конвертируется далее в антигены А.
111
Присутствие Н-, А- и В-трансфераз в сыворотке крови и на эритроцитах устанавливают с помощью специфических антител, которые нередко образуются после трансплантации органов (Eiz-Vesper и соавт. [96]).
Н. В. Бовин и др. (1990) создали искусственные А- и В-субстанции биохимическим синтезом, однако они, несмотря на их структурное сходство с естественным группоспецифическим веществам, не нашли применения, поскольку их адсорбционная активность в отношении α- и β-изогемаагглютининов была низкая.
Молекулярная генетика систем АВО и Н
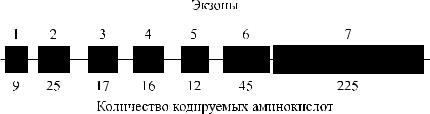
Как уже указывалось выше, серологически определяемые антигены А, В и Н не являются непосредственными продуктами генов. Гены А, В и Н контролируют синтез соответствующих трансфераз. В настоящее время строение генов АВО и Н расшифровано (рис. 3.7). Их удалось клонировать и секвенировать. Расшифрована также аминокислотная последовательность А- и В-трансфераз, которые отличаются друг от друга двумя аминокислотами в позициях 266 и 268 (рис. 3.8).
Рис. 3.7. Строение гена АВО.
MARVLRTLAG |
KPKCHALRPM |
ILFLIMLVLV |
LFGYGVLSPR |
SLMPGSLERG |
50 |
FCMAVREPDH |
LQRVSLPRMV |
YPQPKVLTPC |
RKDVLVVTPW |
LAPIVWEGTF |
100 |
NIDILNEQFR |
LQNTTIGLTV |
FAIKKYVALF |
KLFLETAEKH |
FMVGHRVHYY |
150 |
VFTDQPAAVP |
RVTLGTGRQL |
SVLEVRAYKR |
WQDVSMRRME |
MISDFCERRF |
200 |
LSEVDYLVCV |
DVDMEFRDHV |
GVEILTPLFG |
TLHPGFYGSS |
REAFTYERRP |
250 |
QSQAYIPKDE *GDFYYLGGFF |
GGSVQEVQRL |
TRACHQAMMV |
DQANCIEAVW |
300 |
|
HDESHLNKYL |
**GDFYYMGAFF |
PEYLWDQQLL |
GWPAVLRKLR |
FTAVPKNHQA |
350 |
LRHKPTKVLS |
|||||
VRNP |
|
|
|
|
354 |
Рис. 3.8. Аминокислотная последовательность А-* и B-трансферазы**.
Гены АВО содержат по 1062 пары оснований и кодируют пептиды, состоящие из 354 аминокислот (Reid, Lomas-Francis [186]). Секвенирование генов А и В позволило установить отличающие их последовательности в 7 кодонах, 4 из них (в позициях 176, 235, 266 и 268) могут быть причиной замены аминокислот в трансферазах. Секвенирование гена О1 показало его идентичность с А1 до нуклеотида 261, с которого рамка считывания нарушается с образованием стоп-кодона (Daniels [87]). Такой аллель кодирует синтез пептида, не обладающего какой-либо трансферазной активностью. Возможно также образование
112
нестабильного информационного РНК-транскрипта. Секвенирование гена А 2 выявило делецию одного нуклеотида в кодоне, предшествующем стопкодону в аллеле А1. Делеция приводит к инактивации стоп-кодона и синтезу А-трансферазы, которая содержит 21 аминокислотный остаток дополнительно (Olsson и соавт. [174]). Аллель О, описанный Yamamoto и соавт. [238], в настоящее время получил обозначение О1. Другой аллель О, встречающийся с высокой частотой и получивший обозначение O1v (О1-вариант), также характеризуется делецией в позиции 261. В его присутствии синтез активной трансферазы блокируется (Olsson и соавт. [174]). Еще один вариант аллеля О, обозначенный как О2, не содержит делеции в позиции 261, однако отличается от А1 двумя кодонами, и как результат заменами аминокислотArg 176 Gly и Gly 268Arg. Замена аргинина на глицин приводит к конформационным изменениям молекулы трансферазы и отражается на ее специфической активности.
Синтез АВО-трансфераз контролируют 7 экзонов протяженностью 18 кб, причем на долю экзонов 6 и 7 приходится 77 % генетическго материала (см. рис. 3.7). Делеция G 261 в генах O1 и O1v выявлена в экзоне 6. Замены нуклеотидов, определяющие различия аллелей А, В и О2, локализуются в экзоне 7 (табл. 3.14).
Таблица 3.14
Нуклеотидаяя последовательность в экзонах 6 и 7 локуса АВО, определяющая фенотипические различия
|
|
|
|
|
Позиция нуклеотидов в экзоне |
|
|
|
|
||||||||
|
|
6 |
|
|
|
|
|
|
|
|
7 |
|
|
|
|
|
|
|
2 |
2 |
4 |
5 |
6 |
6 |
6 |
7 |
7 |
7 |
8 |
8 |
8 |
8 |
9 |
1 |
1 |
Локализация |
6 |
9 |
6 |
2 |
4 |
5 |
8 |
0 |
7 |
9 |
0 |
0 |
2 |
7 |
3 |
0 |
0 |
1 |
7 |
7 |
6 |
6 |
7 |
1 |
3 |
1 |
6 |
2 |
3 |
9 |
1 |
0 |
5 |
6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4 |
0 |
Аллель А |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
А101 |
G A C |
C |
T |
C |
G |
G C |
C |
G |
G |
G |
G |
G C |
C |
||||
А102 |
" |
" |
T |
" |
" |
" |
" |
" |
" |
" |
" |
" |
" |
" |
" |
" |
|
А201 |
" |
" |
T |
" |
" |
" |
" |
" |
" |
" |
" |
" |
" |
" |
" |
" |
d |
А301 |
" |
" |
" |
" |
" |
" |
" |
" |
" |
" |
" |
" |
" |
d |
" |
" |
" |
Ах01 |
" " " |
" A |
" " " " " " " " |
" " " " |
|||||||||||||
Цис-АВО1 |
" |
" |
T |
" |
" |
" |
" |
" |
" |
" |
" |
" |
C |
" |
" |
" |
" |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Аллель В |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
В101 |
" |
G |
" |
G |
" |
T |
" |
A |
" |
A |
" |
C |
" |
" |
A |
" |
" |
В301 |
" |
G |
" |
G |
" |
T |
" |
A |
" |
A |
" |
C |
" |
" |
A |
T |
" |
В(А)01 |
" |
G |
" |
G |
" |
" |
" |
" |
" |
A |
" |
C |
" |
" |
A |
" |
" |
Аллель О |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
О01 |
d " " " " |
" " |
" " " " " " " " " " |
||||||||||||||
О02 |
d G " " A " A " T " " " A " " " " |
||||||||||||||||
О03 |
" G " G " " " " " " A " " " " " " |
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
113
Выявлено несколько вставок: гуанин-цитозин (GC boxes), которые находятся выше кодона, инициирующего транскрипцию, и могут играть важную роль в регуляции активности трансфераз. Транскрипция генетической информации зависит от мини-сателлитов – участков размером 4 кб, расположенных выше начального участка считывания. Эксперименты с трансфекцией генов показали, что активность транскрипции гена А по сравнению с В существенно ниже.
Найдены часто встречающиеся аллели, характерные для представителей различных этнических групп. Так, примерно 80 % аллелей А1 среди японцев отличались от аллелей А1 европейцев мутацией, ведущей к замене пролина на лейцин в положении 156. Однако это никак не проявляло себя на фенотипическом уровне (Olsson и соавт. [176–178]). Вместе с тем вставка одного нуклеотида в позиции от 798 до 804 в аллеле А2 приводила к синтезу продукта, не обладающего трансферазной активностью (аллель О 3) (Olsson и соавт. [179]). При сравнении нескольких аллелей установлена их гибридная природа. Образование гибридов, включающих фрагменты двух разных аллелей, авторы объясняют кроссинговером во время мейоза. В большинстве случаев кроссинговер затрагивал экзон 6. При наличии делеции G 261 экзон 7 фенотипически не проявлялся. В тех случаях, когда экзон 6 не содержал указанной делеции, экзон 7 аллеля А1 или О 1 проявлял себя фенотипически как А1-серологическая активность. Экзон 7 аллеля O 1v проявлял себя как А2-серологическая актив-
ность (Ogasawara [170]).
Описана семья, в которой мать имела группу В, ребенок – А, отец – О. На первый взгляд, такие результаты серологического исследования в традиционной интерпретации должны были исключить отцовство. Однако результаты молекулярно-генетического исследования не позволили этого сделать. Секвенирование генов АВО членов данной семьи показало, что у ребенка имелся гибридный ген, экзон 6 которого содержал фрагменты гена В, а экзон 7 – фрагменты гена O 1v. Поскольку экзоны 7 аллелей А1 и O 1v идентичны, а в экзоне 6 указанного гибридного гена, В-O 1v, отсутствовала делеция G 261 (стоп-кодон), на эритроцитах ребенка сформировался антиген А (Olsson и соавт. [177]). Вероятно, этот В-O1 v-гибридный ген с А-трансферазной активностью возник во время мейоза в результате кроссинговера.
В литературе появляется все больше материалов, свидетельствующих о гетерогенности аллелей А, В, Н и Se среди представителей различных рас и этнических групп. Молекулярно-генетические методы позволяют выявлять варианты генов с точковыми мутациями, делециями, гибридными включениями (табл. 3.14, 3.15, 3.16, рис. 3.8), приводящими к формированию стоп-кодонов, инактивации участков сплайсинга (Ogasawara и соавт. [170]). В ряде случаев отмечены эпистатические эффекты в виде аллельного угнетения (Feng и соавт. [101]) или, напротив, аллельного усиления активности генов (Morel и соавт. [162], Ogasawara и соавт. [171]). Структурные особенности генов часто не проявляют себя и лишь в редких случаях приводят к появлению необычных
114
фенотипов с отсутствием или ослаблением экспрессии антигенов А, В и Н [81, 83, 98, 100, 106, 113–115, 124, 126, 131, 141–143, 166, 173, 174, 176–179, 183, 198, 209–211, 220, 222, 242, 248, 249].
Степень гомологии генов АВО человека и высших обезьян достигает 95 %. Это позволяет полагать, что гены АВО возникли не менее 13 миллионов лет назад, то есть значительно раньше разделения высших приматов на отдельные виды: шимпанзе, гориллы, орангутанги и др. (Roubinet и соавт. [190]). Считается, что гены АВО в эволюционном плане высококонсервативны. Эволюционный возраст терминальных углеводных группировок, определяющих групповую принадлежность, вероятно, исчисляется миллиардами лет.
|
|
|
|
|
Таблица 3.15 |
|
|
|
|
Аллели, ассоциированные со слабой экспрессией А |
|||
|
|
|
|
|||
Фенотип |
Аллель |
Замена нуклеотидов |
Замена аминокислот |
|||
|
|
|
|
|||
А3 |
A3-1 |
G 871A |
Asp 291Asn |
|||
А3 |
A3-2 |
G 829A, delC1060 5′ участок |
Val 277 Met, Pro 354 FS |
|||
сплайсинга в интроне 6 |
||||||
|
|
|
|
|
|
|
A |
finn |
Ax-1 |
T 646A |
Phe 216 Ile |
||
|
|
|
|
|||
|
|
|
|
|
|
|
Ax |
Ax-2, -4, -5; |
T646A,G681A,C771T,G829A |
Phe 216 Ile, Val 277 Met |
|||
A1-O1v |
||||||
A |
x |
Ax-3 |
A297 G, T 646A, G 681A |
Phe 216 Ile, Val 277 Met |
||
|
|
|
|
|
||
|
|
|
|
|
||
A |
x |
B-O1v-гибрид |
C 771 T, G 829A |
Phe 216 Ile |
||
|
|
|
|
|
||
|
|
|
|
|
||
A |
x |
Ax-6 |
G 996A |
Thr 332 Стоп |
||
|
|
|
|
|
||
|
|
|
|
|||
Ael |
Ael-1; A*109 |
+ G G798-804 |
Phe 269, FS |
|||
A |
el |
Ael-2; A*110 |
C 467 T, G 829A |
Pro 156 Leu, Phe 216 Ile |
||
|
|
|
|
|
||
|
|
|
|
|
||
A |
w |
Aw-1 |
C 407 T, С 467 T, del C1060 |
Thr 136 Met, Pro 156 Leu |
||
|
|
|
|
|
||
|
|
|
|
|
|
|
Aw |
Aw-2 |
C 350 G, C 467 T, del C 1060 |
Pro 156 Leu, Gly 177Ala, |
|||
Pro 354 FS |
||||||
A |
|
Aw-3 |
G 203 C, C 467 T, del C 1060 |
Arg 68 Thr, Pro 156 Leu, |
||
w |
Pro 354 FS |
|||||
|
|
|
|
|||
|
|
|
|
|
||
A |
w |
Aw-4 |
C 712 T |
Arg 241 Trp |
||
|
|
|
|
|
||
|
|
|
|
|
||
A |
w |
Aw-5 |
A965 G |
Glu 322 Gly |
||
|
|
|
|
|
||
|
|
|
|
|||
Примечание. del – делеция, FS – изменение рамки считывания |
||||||
115