Материал: Донсков С.И., Мороков В.А. Группы крови человека. Руководство по иммуносерологии
эритроцитов специально подобранными трансферазами (α-галактозилазой, экстрагированной из Trichomonas foetus) приводила к появлению Н-активности с одновременной утратой антигена В. Далее эти эритроциты (Н + В −) удавалось конвертировать вAh (А + Н −) добавлением А-трансферазы [195].
Эритроциты лиц Oh Reunion выделителей не агглютинируются большинством анти-Н-сывороток и лишь иногда дают слабоположительные реакции с высокоактивными анти-Н-антителами, присутствующими в сыворотках крови некоторых лиц Oh, или с другими анти-Н-реагентами. Эритроциты таких лиц обычно не реагируют с антителами анти-А и анти-В, однако клетки некоторых индивидов Oh А выделителей могут вести себя в серологических тестах как Ах, демонстрируя слабоположительные реакции с высокоактивными антителами анти-А,В лиц О. Подобные слабые варианты антигена
Вописаны у лиц Oh В.
Всекретах лиц Oh Reunion выделителей содержание вещества Н в норме, субстанции А и В также присутствуют, если данные лица имеют гены А и
В. Сыворотки крови лиц Oh Reunion почти всегда содержат слабые холодовые анти-Н-подобные антитела. Их активность не ингибируется слюной выделителей групповых субстанций, они не реагируют с эритроцитами О новорожденных. Полагают, что эти антитела имеют специфичность анти-HI.
Индивиды, отнесенные к Н-дефицитному типу, найдены среди представителей различных этнических групп – жителей Индии, Европы, Японии, Китая, Юго-Восточной Азии, Среднего Востока, коренных жителей Америки. Их выявляли с частотой от 1 на 5000 среди жителей Таиланда до 1 на 8000 – 1 на 16 000 среди китайцев.
Hm
Еще одна серия Н-дефицитных фенотипов характеризуется следовым количеством или полным отсутствием антигенов А, В и Н на эритроцитах и одновременно нормальным количеством соответствующих группоспецифических субстанций в секретах. Указанные фенотипы обозначены как Om h, Am h и Bm h. В настоящее время их называют Oh, Ah, Bh выделителями. Угнетение синтеза вещества Н у лиц Hm не столь выражено, как у лиц с фенотипами Бомбей, параБомбей и Реюнион. Эритроциты Hm слабо реагируют с анти-Н-антителами, а строма эритроцитов и слюна таких лиц содержит вещество Н в норме.
Остается неясным: имеет ли указанный фенотип какие-либо селективные преимущества в регионе Индийского океана, где он в основном встречается?
Взаимодействие локуса Hh с генами секреции Se и se
Сочетание гена Н с геном Se приводит к тому, что на эритроцитах присутствует вещество Н одновременно с А и / или В в зависимости от того, какой аллель АВО унаследован (табл. 3.13). Если аллель Se по наследству не передается, то вещества А, В и Н в секретах практически отсутствуют (обычные невыделители).
106
Гомозиготные комбинации генов h и se проявляются как Н-дефицит и невыделительство.
Другие варианты гена h способны кодировать синтез вещества Н в небольшом количестве. Последнее преобразуется нормально функционирующими А- и В-трансферазами в небольшое количество антигенов А и В, которые можно выявить с помощью адсорбции – элюции.
Таблица 3.13
Распределение группоспецифических веществ А, В и Н у лиц с Н-дефицитом
Фенотип |
Гены |
АВН-вещества |
АВН-вещества |
|
на эритроцитах |
в слюне |
|||
|
|
|||
|
|
|
|
|
Выделитель АВН |
H, Se, (ABO) |
Н (А или В) |
Н (А или В) |
|
|
|
|
|
|
Невыделитель АВН |
H, sese, (ABO) |
То же |
нет |
|
|
|
|
|
|
Oh Indian тип 1 |
hh, sese, (ABO) |
нет |
То же |
|
Oh Indian вариант |
hh, sese, (ABO) |
Следы Н, А и / или В |
" |
|
Oh Reunion типAh и Bh |
hh, sese, (ABO) |
То же |
" |
|
Н-дефицитный выделитель ОО |
hh, Se, OO |
Следы Н |
Н |
|
|
|
|
|
|
Н-дефицитный выделитель |
hh, Se,A или B |
Следы А или В |
Н, А или В |
|
с наличием генов А или В |
||||
|
|
|
||
|
|
|
|
Как указывалось выше, у гомозигот hh при отсутствии гена Se вещества Н, А и В в секретах отсутствуют. Эти лица способны образовывать анти-Н-антитела.
При отсутствии гена Н (комбинации: hh, Se, OO; hh, Se, A или B) синтеза соответствующего группоспецифического вещества на эритроцитах не происходит. В то же время в плазме крови и секретах вещество Н определяется, поскольку Se- специфическая трансфераза присоединяет L-фукозу к цепям-предшественникам типов 1 и 3. Некоторое количество этой Н-субстанции адсорбируется на эритроцитах, чем и можно объяснить слабоположительные реакции эритроцитов с анти- Н-антителами у обладателей Н-дефицитного фенотипа.
Синтезируемые за счет других трансфераз А- и В-подобные вещества также могут быть адсорбированы эритроцитами и их небольшое количество может быть обнаружено на указанных клетках.
Наилучшим образом возникновение различных Н-дефицитных фенотипов (Бомбей, пара-Бомбей, Реюнион, Hm) объясняет концепция взаимодействия генных локусов H / h и Se / se. Теоретически у родителей Hh, Sese × Hh, Sese возможно рождение детей как с фенотипом Бомбей (hh, sese), так и другими H-дефицитными фенотипами (hh, Sese; hh, SeSe) с наличием выделительства. В действительности такие сочетания генов чрезвычайно редки. Во-первых, аллель h крайне редок. Во-вторых, гены указанных локусов настолько тесно сцеплены между собой, что вероятность рекомбинаций между ними очень мала.
С помощью молекулярно-генетических методов идентифицировано несколь-
107
ко вариантов аллеля h, однако неясно, отличаются ли они между собой в функциональном отношении. Некоторые из указанных аллелей способны кодировать синтез незначительного количества вещества Н. Несмотря на многочисленные доказательства независимости локусов Hh и Sese друг от друга, кодируемые ими фукозил-трансферазы обладают перекрестной способностью к гликозилированию соответствующих исходных субстратов. Так, Н-специфическая трансфераза, присоединяющая L-фукозу к цепям типов 2 и 4, проявляет тропность в отношении цепей типов 1 и 3. Соответственно Se-специфическая трансфераза, гликозилирующая цепи типов 1 и 3, захватывает в этот процесс цепи типов 2 и 4. Такая перекрестная активность в ряде случаев лежит в основе следовой экспрессии антигенов А, В и Н на эритроцитах.
Антигены А, В и Н, адсорбированные из плазмы
Большая часть антигенных веществ А, В и Н синтезируется в процессе эритрогенеза за счет активности соответствующих трансфераз. Вместе с тем некотороее количество указанных группоспецифических субстанций эритроциты адсорбируют из плазмы. Гликосфинголипиды, присутствующие в плазме и несущие вещества Н, А или В, могут встраиваться в мембрану эритроцитов.
Renton и Hancock в 1962 г. обнаружили, что эритроциты группы О, перелитые реципиенту с группой А, приобретают А-антиген. Указанные эритроциты реагировали с антителами анти-А,В и лектином анти-А1 из Dolichos biflorus. С сывороткой анти-А лиц группы В эти эритроциты не реагировали. Авторы установили, что антитела анти-А,В взаимодействуют с детерминантами, расположенными на цепях типа 1 и 2, в то время как лектин распознает антигенную детерминанту, локализованную на цепях типа 2. Подобную картину наблюдали в экспериментах с эритроцитами Oh. После контакта с гликолипидной фракцией плазмы лиц О Le(a −b −) выделителей эритроциты Oh приобретали способность агглютинироваться антителами против цепей 1Н-типа, но не агглютинировались анти-Н-антителами, присутствующими у лиц Oh, а также анти-Н-лектином из Ulex europaeus. Очевидно, что антитела анти-Н, имеющиеся у лиц Oh, подобно анти-Н-лектину из Ulex europaeus, распознают А-антигенную детерминанту на цепях 2Н-типа.
Лимфоциты приобретают антигены Н, А и В из плазмы. Эти антигены встраиваются в мембрану химическим путем в виде гликосфинголипидов, вырабатываемых секреторными клетками. Существует и другое суждение: все групповые АВО-антигенные детерминанты, присутствующие в этх клетках, пассивно адсорбированы ими из плазмы. Изогемагглютинины, нередко содержащиеся в типирующих анти-HLA-сыворотках, могут реагировать с АВО-антигенами лимфоцитов, адсорбированными из плазмы, и искажать результаты лимфоцитотоксического теста.
На тромбоцитах антигены Н, А и В появляются за счет не только адсорбции указанных субстанций из плазмы, но и собственного синтеза. Тромбоциты
108
лиц А2 несут меньше антигена А по сравнению с людьми, имеющими подгруппу А1.
Биохимия антигенов АВО и Н
После открытия групповых антигенов возникла проблема установления их структуры. Задачей иммунохимиков в области групп крови человека являлось выделить и охарактеризовать структуры, ответственные за специфические свойства веществ, обладающих антигенной активностью, и объяснить, почему они независимы в серологических реакциях.
Выделение антигенов А и В из эритроцитов оказалось непростой задачей. На эритроцитах и других клетках они представлены в водонерастворимой форме. Их удалось выделить экстракцией этанолом. Однако вскоре выяснилось, что эти веществасодержатсявомногихорганахитканяхорганизмачеловека,приэтомонирастворимы в воде. Для их выделения к 5 г различных тканей добавляли 25 мл воды, экстракт кипятили в течение 10 мин и затем центрифугировали. Полученный осадок растворяли в 2,5 мл изотонического раствора натрия хлорида. О присутствии субстанций А и В судили по способности экстрактов угнетать активность анти-А-
и / илианти-В-антител(FreidenreichиHartmann,1938).
Эти и другие подобные исследования позволили установить, что наибольшее количество вещества А и В содержится в секреторных тканях (слизистая оболочка желудка, слюнные железы, жидкость кист яичников) и в меконии. Эти же вещества были выделены из стенок желудков лошадей, коров и свиней. Процедуру чаще выполняли замораживанием – оттаиванием экстрактов с последующим растворением высушенного осадка в охлажденном 90 % растворе фенола. Фракция, не поддававшаяся растворению, обладала наибольшей антигенной активностью. Высокой степени очистки удавалось добиться ультрацентрифугированием и использованием органических растворителей, например этанола. Оказалось, что по своей природе группоспецифические вещества А, В и Н являются мукополисахаридами, содержащими приблизительно 85 % углеводов и 15 % белков. Мягкий кислотный гидролиз приводил к исчезновению специфической активности субстрата. При этом высвобождались сахара. Изучение структуры полисахаридов клеточных мембран бактерий подтвердило их антигенные различия, обусловленные именно присутствием тех или иных терминальных углеводных группировок.
Существенный прогресс в изучение природы веществ А, В и Н внесли работы Watkins’a и Morgan’a, показавших присутствие анти-Н-подобных агглютининов в сыворотке угря. Последние вызывали агглютинацию эритроцитов человека группы О. Их активность ингибировалась L-фукозой. При последующих исследованиях обнаружено, что способность анти-А-лектинов агглютинировать эритроциты А устраняется добавлением в них N-ацетил-D-галактозамина. Анти-В-антитела нейтрализовались D-галактозой соответственно. Эти результаты были подтверждены прииспользованииэкзогликозидаз,выделенныхизTrichomonasfoetusиClostridium
109
welchii. Указанные ферменты разрушали вещества А, В и Н. В то же время актив- ностьэтихферментовустранялиN-ацетил-D-галактозамин,D-галактозаиL-фукоза соответственно,чтоуказывалонахимическуюприродугрупповоговещества.
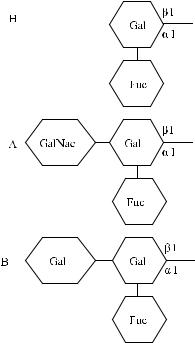
В настоящее время химическая структура групповых веществ хорошо изуче-
на (рис. 3.6).
Рис. 3.6. Структура антигенных детерминант Н, А и В.
Антигены систем АВО и Н представляют собой олигосахаридные цепи, связанные с полипептидами (гликопротеины) или церамидами (гликосфинголипиды).
Выделяют 2 класса олигосахаридных цепей, которые экспрессируют АВНантигены. Первый из них представлен N-гликанами – разветвленными структурами, связанными через аминогруппы аспарагина, и N-ацетилглюкозамин. Второй класс представлен О-гликанами, имеющими простую или сложную структуру, связывание в них происходит через гидроксильные группы серина или треонина также через N-ацетилглюкозамин.
Гликосфинголипиды (углеводные цепи, присоединенные к церамиду) подразделяют в зависимости от биохимической природы на глобозиды, лактозиды и ганглиозиды.
Основная масса антигенов Н, А и В организма представлена гликопротеинами, доля гликосфинголипидов существенно меньше.
110