действию комплемента и в большей степени подвержены гемолизу (Bobolis и
соавт. [3], Telen и соавт. [22]).
Описаны 2 больных дизэритропоэтической анемией, эритроциты которых содержали слабый антиген JMH (Bobolis и соавт. [2]).
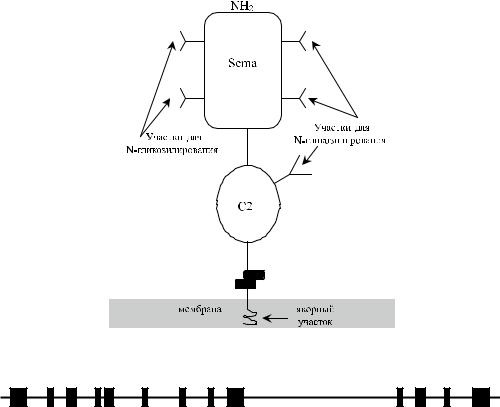
Секретируемые и мембранные семафорины выполняют функцию сигнальных белков (белков наведения), способствующих росту аксонов нервной клетки в нужном направлении (Bron и соавт. [5], Kolodkin и соавт. [13]), участвуют в межклеточных взаимодействиях (Tamagnone, Comoglio [21]).
Плексины – основные рецепторы семафоринов – участвуют в регуляции деятельности эндокринной, иммунной, сердечно-сосудистой, желудочно-кишечной и других важнейших систем организма (Yazdani, Terman [26]).
Гликопротеин CDw108 экспрессирован преимущественно на активированных лимфоцитах и содержит аминокислотную последовательностьArg-Gly-Asp (267–269), характерную для молекул клеточной адгезии. Большое количество РНК-транскриптов SEMA7A выявлено в плаценте, тестикулах, селезенке, низкое – в головном мозге и тимусе (Lange и соавт. [14],Yamada и соавт. [25]).
Утрата антигенов JMH (и обретение фенотипа JMH −) in vivo может быть обусловлена действием внутриклеточных протеаз (Bobolis, Telen [4]). Инкубация эритроцитов при 37 оС в течение 45 мин приводила к появлению в супернатанте серологически активного JMH-протеина с мол. массой 67 кДа. Отделение протеина тормозилось ингибиторами протеаз. Высвобождение протеина JMH из изолированных мембран происходило только при добавлении цитоплазмы или нейтрофилов (Bobolis, Telen [4]).
Список литературы
1.Baldwin M.L., Ness P.M., Barasso C. et al. In vivo studies of the long term 51Cr red cell survivalofserologicallyincompatibleredcellunits//Transfusion.–1985.–V.25.–P.34–38.
2.Bobolis K.A., Lande W.M., Telen M.J. Markedly weakened expression of JMH in a kindred with congenital hemolytic anemia [Abstract] // Transfusion. – 1991. – V. 31. – 46S.
3.Bobolis K.A., Moulds J.J., Telen M.J. Isolation of the JMH antigen on a novel phosphtidylinositol-linked human membrane protein // Blood. – 1992. – V. 79. – P. 1574– 1581.
4.Bobolis K.A., Telen M.J. Biochemical study of possible mechanisms of acquired loss of JMH antigen expression [Abstract] // Transfusion. – 1991. – V. 31. – 46S.
5.Bron R., Vermeren M., Kokot N. et al. Boundary cap cells constrain spinal motor neuron somal migration at motor exit points by a semaphorin-plexin mechanism // Neural Dev. – 2007. – V. 2. – P. 21.
6.Daniels G.L., Flegel W.A., Fletcher A. et al. International Society of Blood Transfusion committee on terminology foe red cell surface antigens: Cape Town report // Vox Sang. – 2007. – V. 92. – P. 250–253.
7.Daniels G.L., Knowles R.W.Amonoclonal antibody to the high frequency red cell antigen JMH // J. Immunogenet. – 1982. – V. 9. – P. 57–59.
8.Daniels G.L., Knowles R.W. Further analysis of the monoclonal antibody H8 demonstrating a JMH-like specificity // J. Immunogenet. – 1983. – V. 10. – P. 257–258.
9.Geisland J., Corgan M., Hillard B. An example of anti-JMH with characteristics of a clinically significant antibody. //Immunohematology. – 1990. – V. 6. – P.9-11.