Материал: Донсков С.И., Мороков В.А. Группы крови человека. Руководство по иммуносерологии
Феномен перекрестного реагирования сывороток Оαβ(I) впервые описали Landsteiner и Witt в 1926 г. [134] как парадокс, нарушающий стройную концепцию системы АВО, постулирующую 2 агглютиногена – А и В, и два агглютинина – α и β. Вопреки ожиданиям исследователей элюаты, полученные при адсорбции сывороток Оαβ(I) эритроцитами А, реагировали не только с эритроцитами А, но и с эритроцитами В. Такой же результат наблюдали после адсорбции сывороток Оαβ(I) эритроцитами В: элюат реагировал с эритроцитами В и А.
При адсорбции смеси сывороток Аβ(II) и Вα(III) перекрестного реагирования антител не происходило: элюат с эритроцитов А реагировал только с эритроцитами А, элюат с эритроцитов В – только с эритроцитами В (Wiener и соавт. [229]).
Адсорбционные пробы с сывороткой Оαβ(I), а именно возможность удалить с помощью эритроцитов А агглютинины β, а с помощью эритроцитов В – агглютинины α, указывают на то, что антигены А и В содержат общий компонент, который и является, по Винеру, антигеном С.
Весьма убедительными казались доводы оппонентов, отрицавших существо-
вание антигена С. Так, Dodd, Lincoln, Boorman [91, 146], Bird [80] полагали,
что анти-С-антитела представляют собой химически сшитые, несепарируемые, αβ-молекулы, способные реагировать с антигенами А и В. Таким образом, антиген С на эритроцитах А и В – это лишь видимость.
Другиевидныеисследователи(RaceиSanger[184],Доссе[25],П.Н.Косяков[35], Прокоп [56]) также указывали, что α- и β-агглютинины лиц группы О(I) не являются простойсмесью.Логичнодопустить,чтовпроцессесинтезаα-агглютининовиодно- временноβ-агглютининовкакая-точастьизнихможетбытьсобранакакαβ-антитела.
Важным аргументом против существования антигена С явился также тот факт, что до настоящего времени не найдены индивиды, в эритроцитах которых антиген С присутствует в чистом виде без антигенов А и В.
По мнению Wiener [229], некоторые образцы эритроцитов Аx (А4,А5) как раз и являются носителями антигена С в чистом виде. Эти эритроциты не содержат антигена В, а количество антигена А в них крайне мало. Особенностью этих эритроцитов является то, что они не реагируют с сыворотками Вα(III), но реагируют с сыворотками Оαβ(I), так как сыворотки Вα(III) содержат только α-агглютинины, асывороткиОαβ(I)нарядусα-агглютининамиимеютанти-С-антитела.Болеетого, у лиц Аx в сыворотке крови иногда содержатся α1-экстраагглютинины, поэтому нельзяисключать,чтоантигеныА4 иА5таковыминасамомделенеявляются,т.е. не относятся к группе А, а представляют собой антиген С.
Race и Sanger [184] не разделяли точку зрения Винера, указывая на то, что некоторые сыворотки Вα(III) слабо, но все-таки реагируют с эритроцитами А4, А5 [184]. Кроме того, в слюне людей А4, А5 находят растворимый антиген А, нейтрализующий α-агглютинины сывороток Вα(III), что свидетельствует о принадлежности антигенов А4 и А5 к группе А.
Дискуссия относительно существования антигена С в системе АВО, продолжавшаяся до 1970-х годов, в последующие годы не возобновлялась в связи с
81
отсутствием новых данных, а также некоторым скептическим отношением иммуносерологов к этому вопросу, не представляющему, по их мнению, большого значения для практики переливания крови.
На наш взгляд, перекрестно реагирующие антитела имеют значение в трансфузиологии. Их присутствие у реципиентов, по-видимому, усугубляет тяжесть посттрансфузионных осложнений. Об этом свидетельствует то обстоятельство, что среди реципиентов О(I), которым перелили кровь другой группы, осложнения протекают более тяжело и риск летального исхода выше, чем у реципиентов A(II) и B(III), которым перелили кровь другой группы. Однако соответствующие иммуносерологические исследования, подтверждающие или опровергающие высказанное положение, не были проведены.
Вместе с тем в литературе накапливались сведения о том, что перекрестно реагирующие антитела, а стало быть и антиген С, имеют значение в акушерстве.
Rosenfield в 1953 г. [188, 189], исследуя групповые антитела у новорожденных и их матерей, обратил внимание на 2 обстоятельства: 1 – если мать и плод имели группу О(I), то у новорожденных α- и β-агглютинины в сыворотке крови присутствовали чаще, чем в случаях, когда мать и плод имели группу A(II) или оба относились к группе B(III); 2 – у женщин с О(I) группой крови ГБН развивалась чаще, чем у рожениц, имевших другие группы крови.
Unger и Wiener [213], Wiener и Wexler [231], подтвердив данные Rosenfield,
показали, что у новорожденных О(I) от матерей О(I) через месяц после рождения перекрестно реагирующие αβ-антитела исчезали, что свидетельствовало об их материнском происхождении.
Kochwa и Rosenfield установили, что перекрестно реагирующие антитела анти-А,В относятся к глобулинам 7Sγ2, которые легкопроникают через плаценту, поскольку размер их молекул меньше, чем у других антител.
Разделяя взгляды Винера, мы предприняли попытку найти подтверждение его концепции, а именно того, что перекрестные реакции в системе АВО обусловлены антигеном С эритроцитов, а не комбинированными αβ-антителами.
Дляначалапредставлялосьцелесообразнымполучитьсобственныеданныеохарактереперекрестныхреакций,свойственныхсывороткамОαβ(I)[20,21,23,24,47].
Материал и методы. Исследовали сыворотки крови 100 доноров, имеющих группу крови О(I), мужчин и женщин. Каждую сыворотку титровали с эритроцитами А и В до и после адсорбции эритроцитами А, В и О. Титр сывороток устанавливали кратным их разведения 0,9 % NaCl. В качестве контроля первые 3 разведения сыворотки тестировалиэритроцитамиО(I).Адсорбцию антител проводили при комнатнойтемпературе 20–22 оСвтечение 1 ч в соотношении2 объемасыворотки +1 объем плотно упакованного гомогенизированного осадка трижды отмытых эритроцитов. Устойчивость антител к унитиолу (2,3-димеркаптопропансульфонат натрия – CH2SH-CH-SH-CH2SO3Na) оценивали титрованием сыворотки после добавления равного объема 1,25 % раствора этого реактива и инкубации полученной смеси в течение 2 ч при температуре 37 оС. Температурную устойчивость антител исследовали посредством прогревания сывороток при температуре 56 и 70 оС в течение 1 ч и 10 мин соответственно.
82
Из 100 исследованных сывороток группы О(I) 52 не содержали перекрестно реагирующих антител. При уменьшении титра α-антител на 4–8 ступеней после адсорбции этих сывороток эритроцитами А β-антитела оставались в том же титре, что и до адсорбции. Аналогичный результат наблюдали после адсорбции этих сывороток эритроцитами В. Титр β-антител снижался на 4–8 ступеней, в то время как титр α-антител оставался прежним (табл. 3.7).
Таким образом, 52 % сывороток Оαβ(I) не обладали способностью к перекрестному реагированию, 48 % сывороток содержали перекрестно реагирующие антитела. Перекрестное реагирование было симметричным и асимметричным, в частности, 20 из 48 сывороток показали симметричное перекрестное реагирование, т. е. независимо от того, какими эритроцитами (А или В) проводили их адсорбцию, титр α- и β-агглютининов снижался пропорционально.
Другие 28 сывороток показали асимметричное перекрестное реагирование. В 15 сывороткахпослеадсорбцииэритроцитамиАснижалсятитробоихагглютининов(α и β), а после адсорбции эритроцитами В убывал титр только β-агглютининов, титр α-антител оставался прежним. В 13 сыворотках, наоборот, снижение титра обоих агглютининовнаблюдалипослеадсорбцииэритроцитамиВ.Адсорбцияэритроцитами Аснижалатитрα-агглютининов,ноневлияланатитрβ-антител.Такимобразом,пе- рекрестное реагирование по своему характеру может быть симметричным, асимме- тричнымвсторонуА-антигенаиасимметричнымвсторонуВ-антигена.
|
|
|
|
|
|
|
|
|
|
|
|
|
Таблица 3.7 |
||
|
Варианты перекрестной адсорбции сывороток Оαβ(I) |
|
|
|
|||||||||||
Категория |
Серия |
Титр |
|
|
Титр после адсорбции эритроцитами |
|
|||||||||
до адсорбции |
|
|
A(II) |
|
B(III) |
|
|
О(I) |
|
||||||
сыворотки |
сыворотки |
с эритроцитами |
с эритроцитами |
с эритроцитами |
с эритроцитами |
||||||||||
|
|
A |
В |
О |
А |
В |
О |
А |
В |
О |
А |
|
В |
О |
|
|
399 |
1 : 256 |
1 : 256 |
|
0 |
1 : 2 |
1 : 256 |
0 |
1 : 256 |
0 |
0 |
1 : 2 56 |
|
1 : 256 |
0 |
I |
626 |
1 : 32 |
1 : 8 |
|
0 |
0 |
1 : 8 |
0 |
1 : 32 |
0 |
0 |
1 : 32 |
|
1 : 8 |
0 |
997 |
1 : 256 |
1 : 128 |
|
0 |
0 |
1 : 128 |
0 |
1 : 256 |
0 |
0 |
1 : 256 |
|
1 : 128 |
0 |
|
|
|
|
|||||||||||||
|
846 |
1 : 64 |
1 : 32 |
|
0 |
0 |
1 : 32 |
0 |
1 : 64 |
0 |
0 |
1 : 64 |
|
1 : 32 |
0 |
|
129 |
1 : 128 |
1 : 128 |
|
0 |
1:2 |
1 : 32 |
0 |
1 : 64 |
0 |
0 |
1 : 128 |
|
1 : 128 |
0 |
II |
272 |
1 : 256 |
1 : 128 |
|
0 |
0 |
1 : 64 |
0 |
1 : 64 |
0 |
0 |
1 : 256 |
|
1 : 128 |
0 |
397 |
1 : 256 |
1 : 64 |
|
0 |
0 |
1 : 32 |
0 |
1 : 32 |
1 : 2 |
0 |
1 : 256 |
|
1 : 64 |
0 |
|
|
|
|
|||||||||||||
|
488 |
1 : 256 |
1 : 32 |
|
0 |
0 |
1 : 8 |
0 |
1 : 32 |
0 |
0 |
1 : 256 |
|
1 : 32 |
0 |
|
127 |
1: 256 |
1 : 128 |
|
0 |
1 : 2 |
1 : 64 |
0 |
1 : 256 |
1 : 2 |
0 |
1 : 256 |
|
1 : 128 |
0 |
III |
855 |
1 : 128 |
1 : 64 |
|
0 |
0 |
1 : 32 |
0 |
1 : 128 |
0 |
0 |
1 : 128 |
|
1 : 64 |
0 |
505 |
1 : 256 |
1 : 128 |
|
0 |
0 |
1 : 64 |
0 |
1 : 128 |
0 |
0 |
1 : 256 |
|
1 : 128 |
0 |
|
|
|
|
|||||||||||||
|
705 |
1 : 512 |
1 : 256 |
|
0 |
0 |
1 : 128 |
0 |
1 : 512 |
0 |
0 |
1 : 512 |
|
1 : 256 |
0 |
|
400 |
1 : 128 |
1 : 128 |
|
0 |
0 |
1 : 128 |
0 |
1 : 64 |
0 |
0 |
1 : 128 |
|
1 : 128 |
0 |
IV |
424 |
1 : 256 |
1 : 64 |
|
0 |
0 |
1 : 64 |
0 |
1 : 128 |
0 |
0 |
1 : 256 |
|
1 : 64 |
0 |
454 |
1 : 256 |
1 : 128 |
|
0 |
1 : 2 |
1 : 128 |
0 |
1 : 128 |
1 : 2 |
0 |
1 : 256 |
|
1 : 128 |
0 |
|
|
|
|
|||||||||||||
|
351 |
1 : 256 |
1 : 128 |
|
0 |
0 |
1 : 128 |
0 |
1 : 128 |
0 |
0 |
1 : 256 |
|
1 : 128 |
0 |
Примечание.КатегорияI–сыворотки,несодержащиеперекрестнореагирующихантител, II – сыворотки, содержащие антитела, дающие симметричные перекрестные реакции, IIIиIV– сыворотки,содержащиеантитела,дающиеасимметричныеперекрестныереакции.
83
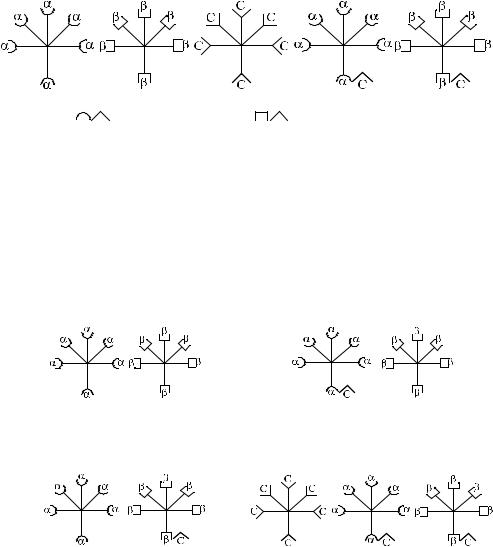
Нанашвзгляд,перекрестноереагированиеобусловленонеαβ-агглютининами, как полагала Dodd и соавт., а антителами αС, βС, что в большей мере согласуется с концепцией Винера, а также полученными нами данными. На рис. 3.2 представлена предполагаемая модель этих антител.
1 |
2 |
3 |
4 |
5 |
|
– антиген А по Винеру |
|
– антиген В по Винеру |
|
Рис. 3.2. Возможные варианты изогемагглютининов в сыворотке Оαβ(I).
1 – α-изогемагглютинины; 2 – β-изогемагглютинины, 3 – С-изогемагглютинины; 4 – αС-изогемагглютинины; 5 – βС-изогемагглютинины.
Представленная структурная модель изогемагглютининов позволяет объяснить характер перекрестного реагирования сывороток Оαβ(I). Сыворотки, не вызывающие перекрестных реакций, содержат только α- и β-антитела (рис. 3.3, п. 1). Сыворотки, вызывающие симметричные перекрестные реакции, содержат, помимо α- и β-антител, антитела αС и βС или анти-С-антитела (рис. 3.3, п. 4). Сыворотки, вызывающие асимметричные перекрестные реакции, содержат α, β и одну из разновидностей антител – αС или βС (рис. 3.3, п. 2, 3).
1.Перекрестная реакция отсутствует 2. Асимметричная перекрестная реакция
(α-агглютинины не могут адсорбироваться на эритроцитах В, β-агглютинины не могут адсорбироваться на эритроцитах А).
(α-агглютинины могут адсорбироваться на эритроцитах В за счет присутствующего на них антигена С).
3.Асимметричная перекрестная реакция 4. Симметричная перекрестная реакция
(β-агглютинины могут адсорбироваться на эри- |
(1-й вариант: β-агглютинины адсорбируются на |
троцитах А за счет присутствующего на них ан- |
эритроцитах А, α-агглютинины адсорбируются |
тигена С). |
на эритроцитах В за счет рецептора анти-С, 2-й |
|
вариант: анти-С-агглютинины адсорбируются |
|
на эритроцитах А и В). |
Рис. 3.3. Комбинации агглютининов, обусловливающие различные варианты перекрестного реагирования.
84
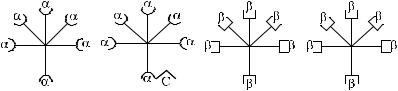
Рассмотрим модель асимметричного реагирования сыворотки, которая содержит агглютинины в сочетании, представленном на рис. 3.4.
Рис. 3.4. Вариант асимметрично реагирующей сыворотки.
Согласно схеме сыворотка содержит 50 % α-агглютининов, 75 % – β. Валентность анти-С реагирует как α и β. При адсорбции такой сыворотки эритро- цитамиАα-агглютининыбудутудаленыполностью.Инымисловами,изсыворот- ки будет удалено 50 % антител. При адсорбции сыворотки эритроцитами В будет удалено 75 % антител (50 % β + 25 % α) за счет рецептора анти-С. В случае, если рецептор анти-С связан не с α-, а с β-агглютининами, асимметричное реагирование проявят эритроциты А, унося на своей поверхности 25 % β-агглютининов.
Какдалеенамипоказано,перекрестнореагирующиеантителатакже,какиммунныеантителаАВОиRh,относятсякклассуIgG,устойчивыкунитиолу,лучшереагируют с отмытыми, чем неотмытыми эритроцитами, в отличие от иммунных (термостабильных)антителявляютсятермолабильными(утрачиваютактивностьпослепрогревания сыворотки при 70 оС в течение 10 мин). В противоположность классическимиммуннымантителамIgG,которыеадсорбируютсянаэритроцитах,ноневызывают их агглютинации, перекрестно реагирующие антитела непосредственно агглютинируют эритроциты в солевой среде. В низкой концентрации они утрачивают способность агглютинировать эритроциты, однако сенсибилизируют их подобно неполнымантителамихорошовыявляютсявнепрямойантиглобулиновойпробеКумбса.
Сыворотки лиц Оαβ(I), очевидно, содержат несколько типов групповых анти- тел:изогемагглютининыαиβIgM,αиβIgG,αСиβСIgG,анти-СIgG,иммунные α и β IgM, иммунные α и β IgG, что закономерно обусловливает перекрестные реакции с различными групповыми антигенами.
Важным аргументом, подтверждающим концепцию Винера о существовании антигена С, явились эксперименты по получению специфических антисыворотк. В частности, при иммунизации мышей эритроцитами А(II) и В(III) нами [21] были полученымоноклональныеантителасоспецифичностьюанти-АВ(МКА-АВ).
Материал и методы. Трехмесячным самкам мышей BALB / с вводили по 0,1 мл отмытых 0,9 % NaCl эритроцитов А(II) или В(III) внутрибрюшинно дважды с интервалом в 7 дней. Третью инъекцию производили внутривенно за 5 дней до слияния спленоцитов с клетками миеломы NS-1. Слияние осуществляли по стандартной методике с использованием 45 % полиэтиленгликоля-1500 «Loba» и 5 % диметилсульфоксида. Эффективность гибридизации составляла 10–5. Клонировали гибридные клетки в среде НАТ без фидера. Антителопродуцирующие клоны отбирали с помощью реакции агглютинации в 96-луночных планшетах с отмытыми эритроцитами О(I), А(II) и В(III).
Класс моноклональных антител устанавливали по флюоресценции сенсибилизи-
85