MTVA |
RPSVPAALPL |
LGELPRLLLL |
VLLCLPAVWG |
-1 |
DCGLPPDVPN AQPALEGRTS FPEDTVITYK CEESFVKIPG EKDSVICLKG |
50 |
SQWSDIEEFC NRSCEVPTRL NSASLKQPYI TQNYFPVGTV VEYECRPGYR |
100 |
REPSLSPKLT CLQNLKWSTA VEFCKKKSCP NPGEIRNGQI DVPGGILFGA |
150 |
TISFSCNTGY RLFGSTSSFC LISGSSVQWS DPLPECREIY CPAPPQIDNG |
200 |
IIQGERDHYG YRQSVTYACN KGFTMIGEHS IYCTVNNDEG EWSGPPPECR |
250 |
GKSLTSKVPP TVQKPTTVNV PTTEVSPTSQ KTTTKTTTPN AQATRSTPVS |
300 |
RTTKHFHETT PNKGSGTTSG |
TTRLLSGHTC |
FTLTGLLGTL |
VTMGLLT |
347 |
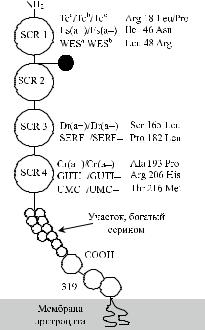
Рис. 21.2. Аминокислотная последовательность DAF.
Клиническое значение
Антитела системы Cromer представлены преимущественно классом IgG (Daniels [15]), встречаются анти-Cr a-антитела IgM (Dickson и соавт. [23]).
Большинство антител IgG относится к субклассу IgG1, однако описаны анти-
тела, относящиеся к субклассам IgG2, IgG3 и IgG4 (Dickson и соавт. [23], Issitt, Anstee [36], Nakache и соавт. [66], Reid и соавт. [79, 80, 83], Sistonen и соавт. [89], McSwain, Robins [59],Anderson и соавт. [1], Byrne и соавт. [7]).
Антитела системы Cromer не имеют существенного значения в трансфузиологии. Опубликованы сообщения о благополучных исходах трансфузий эритроцитов, несовместимых по антигенам этой системы, реципиентам, которые име-
ли антитела анти-Cr a (Smith и соавт. [90], Whitsett, Oxendine [107], Chapman и соавт. [9]) и анти-Tc a (Hoffer и соавт. [30]).
Вместе с тем описаны посттрансфузионные реакции, причиной которых были антитела анти-Cr a (Byrne и соавт. [7]) и анти-Tc a (Kowalski и соавт. [39]). В последнем случае реципиенту перелили шесть доз эритроцитов Tc(a + ), из которых первые три гемолизировались in vivo в день проведения трансфузии.
Попытки оценить клиническое значение антител Cromer с помощью экспериментальных тестов in vivo и in vitro не дали однозначных результатов.
Smith и соавт. [90], Leatherbarrow и соавт. [45] и другие авторы [4, 9, 23, 66, 79, 84, 86, 107,] сделали вывод об отсутствии клинического значения антигенов
иантител этой системы.
Вдругих случаях были получены данные, свидетельствующие о способности антител Cromer уменьшать продолжительность циркуляции перелитых эри-
троцитов [1, 7, 27, 38, 39, 43, 59, 79, 83, 105].
ПривведенииэритроцитовIFC +реципиенту,имевшемуанти-IFC-антитела,отме- чалосьследующее.Водномслучаечерезсуткипослеинъекциивкровотокесохранилось38 %эритроцитов(Walthersисоавт.[105]),вдругомслучаенаблюдалиисчезно- вениеIFC-несовместимыхэритроцитовчерез15минпослевведения(Daniels[15]).
Антитела анти-Tc a IgG1, IgG2 и IgG4 проявляли себя как клинически значимые при испытании в монослое моноцитов. Через 2 года антитела того же лица содержали только IgG2 и IgG4 и не проявляли себя in vitro как клинически зна-
чимые (Anderson и соавт. [1]).
Трансплантация почки от донора Dr(a + ) реципиенту Dr(a −), имевшему анти- Dr a-антитела IgG2 и IgG4, была успешной: приживление трансплантата и его