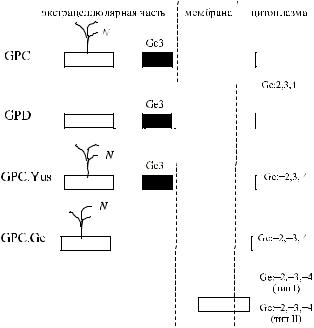
исоавт. [110]). Аллель GPC.Ge кодирует гликофорин-С-подобную структуру, лишенную аминокислот в положениях 36–63. GPC.Ge несколько меньше гликофорина GPC.Yus, поскольку экзон 2 лишен вставки из 27 нуклеотидов, присутствующей в экзоне 3.
Еще один Gerbich-отрицательный фенотип, Ge: −2, −3, −4, получил обозначение Leach. Он является истинно нулевым фенотипом: эритроциты лиц с указанной редкой группой крови лишены всех антигенов системы Gerbich. В литературе имеются описания шести лиц Ge: −2, −3, −4. Все они оказались европеоида-
ми (англичане, американцы) (Anstee и соавт. [5], Daniels и соавт. [30], McShane, Chung [67], Reid и соавт. [98], Rountree и соавт. [107]).
Вдвух семьях удалось показать наследование рецессивных генов, обусловливающих возникновение фенотипа Ge: −2, −3, −4 (Daniels и соавт. [30], Reid и соавт. [98]). На эритроцитах лиц с этой крайне редкой группой крови гликофорины С и D полностью отсутствуют (Anstee и соавт. [5], Daniels и соавт. [30], High и соавт. [39], Reid и соавт. [94, 98]).
Фенотип Leach (Ge: −2, −3, −4) может быть обусловлен разными причинами. У 5 не состоящих в родстве лиц, выявлена делеция экзона 3 и 4 гена GYPC
(High и соавт. [39], Johnson, Daniels [44], Telen и соавт. [121], Winardi и соавт. [129]). В то же время из ретикулоцитов лиц Ge: −2, −3, −4 были выделены обычные транскрипты гена GYPC (Winardi и соавт. [129]).
У другого индивида Ge: −2, −3, −4 выделен полноценный ген GYPC, однако при секвенировании экзона 3 отмечена мутация, образующая стоп-кодон в позиции 56 (Telen и соавт. [121]). При этом не транслировалась большая часть экзона 3 и весь экзон 4. Тем не менее у обследуемых был выявлен небольшой фрагмент с мол. массой 12 кДа, напоминавший терминальную часть гликофоринов С и D (Pinder и соавт. [88]). В связи с этим высказано предположение, что трансляция дефектного гена GYPC может начинаться в других точках.
Имеется несколько сообщений о низкой экспрессии на эритроцитах Ge: −2, −3 антигенов Kell. Степень подавления у индивидов Ge: −2, −3 варьиро-
вала (Anstee и соавт. [5], Daniels и соавт. [25, 30], McShane, Chung [67], Nash
исоавт. [77], Okubo и соавт. [81]). Снижение экспрессии отмечено в отношении часто встречающихся антигенов Kell, в также антигена KEL1. У одного индивида отмечена слабая экспрессия антигена KEL11 (Daniels [25]), выраженность других антигенов соответствовала норме. В 9 из 11 образцов эритроцитов Ge: −2, −3 отмечена слабая выраженность антигенов Kell. В то же время шесть образцов эритроцитов Ge: −2, 3 имели нормальную экспрессию этих антигенов.
У 4 лиц Ge: −2, −3, −4 экспрессия антигенов Kell была слабой (Anstee и соавт. [5], Daniels и соавт. [30], McShane, Chung [67]).
Фенотипическая зависимость антигенов Kell от системы Gerbich реализуется, по-видимому, не на генетическом уровне, поскольку гены, контролирующие синтез вещества Gerbich и вещества Kell, расположены на разных хромосомах и непосредственное влияние их друг на друга исключено.