связаны между собой на хромосоме 6 в регионе III большого комплекса гистосовместимости. Некоторые гаплотипы С4А и HLA показывают высокую степень неравновесного сцепления. Так, гаплотип C4A*Q0 у европеоидов ассоциирован с фенотипом HLA-A1,B8,DR3, у негроидов – с фенотипом HLA-В44,DR2, гаплотип C4В*Q0 чаще встречается у лиц HLA-B5,B12.
Серологический полиморфизм
С помощью реакции нейтрализации специфических антител выявлено три фенотипа Rodgers: Rg +, Rg − и частично ингибирующий Rg + (Longster, Giles [47]), а также четыре фенотипа Chido: Ch +, Ch − и два частично ингибирующих
Ch + (Giles и соавт. [24], Nordhagen и соавт. [63]).
Идентифицированы две разновидности антител анти-Rodgers (анти-Rg1 и анти-Rg2) и три разновидности антител анти-Chido (анти-Ch1, анти-Ch2 и антиCh3), распознающих антигены, имеющие высокую частоту (Giles и соавт. [24, 22, 23]).
Плазма Rg +, полностью ингибирующая анти-Rodgers-антитела, была получена от лиц с фенотипом Rg:1, 2. Плазма Rg −, не ингибирующая указанные антитела, принадлежала индивидам Rg:−1,−2. Частичную ингибицию анти- Rodgers-антител вызывала плазма лиц Rg:1, −2. Фенотип Rg: −1, 2 не найден.
Плазма, ингибирующая активность анти-Chido-антител, получена от лиц Ch:1, 2, 3, плазма, не обладающая такой способностью, – от индивидов
Ch:− 1, −2, −3. Плазма лиц Ch:1, −2, 3, Ch:1, 2, −3 и Ch:1, −2,3 вызывала частич-
ную ингибицию антител анти-Chido. Три последних фенотипа встречаются ред-
ко (Giles и соавт. [38], Skanes и соавт. [85]). Фенотипы Ch: −1, 2, 3 и Ch: −1, −2, 3
не найдены (Yu и соавт. [100]).
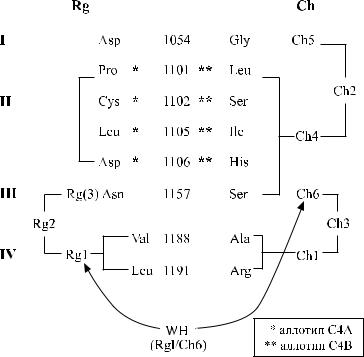
Молекулы С4 экспрессируют антиген Rg1 или Ch1. Оба антигена вместе на эритроцитах не присутствуют.
Частично ингибирующий фенотип Rg:1, −2 находят преимущественно у лиц, наследующих гаплотип С4А*3А*2В*Q0. Отмечена выраженная ассоциация фенотипа Ch:1, −2, 3 с гаплотипом С4В*2, фенотипа Ch:1,2, −3 с гаплотипами С4А*6В*1 и С4А*3В*1. В остальном фенотипы Ch и Rg не коррелируют с алло-
типами С4 (Giles и соавт. [28]).
Серология системы Ch / Rg еще более усложнилась после открытия Giles [27] трех новых часто встречающихся антигенов: Ch4, Ch5 и Ch6. Антитела к этим детерминантам могут быть выявлены только с использованием эритроцитов Ch: −1, −2, −3, нагруженных компонентом С4 различных аллотипов, в том числе с обратной антигенностью Ch / Rg.
Антиген Ch4 свойствен всем С4В-аллотипам, но не встречается при наличии нулевых гаплотипов: C4B*Q0 и C4A*1B*Q0. Последний кодирует антигены Ch1 и Ch3 в отсутствие Rg1 и Rg2.
Антиген Ch5 ассоциирован с Ch2 на протеине С4В, но может присутствовать на С4А1 без Ch2.