существенно снижена (иногда до нуля), но спустя некоторое время после родов восстанавливается [115].
Исследуя особенности возрастной трансформации антигенов Lewis у новорожденных и детей, М.А. Бронникова [3] пришла к ряду важных выводов, которые можно считать классическими, поскольку они в полной мере отражают состояние системы Lewis в этот период онтогенеза:
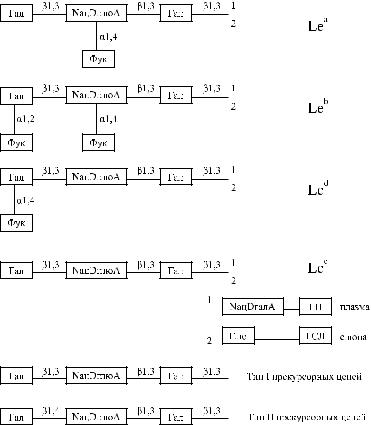
––в сыворотке новорожденных присутствуют оба вещества (Le а и Le b), а на эритроцитах они отсутствуют. Большинство новорожденных имеют фенотип Le(a −b −).
––у детей определяются антигены, отсутствующие у обоих родителей. Большинство детей до года имеют фенотип Le(a +b + ). Антиген Le а присутствует чаще, чем Le b. Lawler и Marshall [140, 141] полагают, наоборот, что Le b чаще.
––фенотип Le(a +b −) и отсутствие секреции АВН в первые годы жизни часто не связаны. Фенотип Le(a +b −) чаще сочетается с группой О(I).
––группа Le(a −b + ) формируется за счет постепенного уменьшения синте-
за Le а в группе Le(a +b + ).
––антигены Lewis окончательно формируются к 5 годам жизни, но возрастная трансформация по годам весьма индивидуальна. Группа
Le(a +b −) формируется раньше, чем Le(a −b + ).
––агглютинабельность эритроцитов новорожденных при воздействии на них сыворотками анти-Le а и анти-Le b выражена в меньшей степени, чем у взрослых, и не усиливается после обработки трипсином, а с
3-летнего возраста усиливается, как и у взрослых.
В эмбриональном и раннем постнатальном периоде, как полагает М.А. Бронникова [3], синтезируются не Lewis-антигены, а их предшественник, который затем трансформируется в Le а или Le b или утрачивает серологическую активность. Редкие случаи фенотипа Le(a +b + ) следует рассматривать как нарушение нормального процесса синтеза антигенов Lewis.
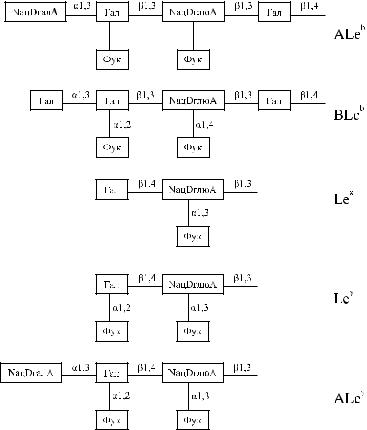
Считается [115], что Se- и Le-генспецифические трансферазы продуцируются в меньшем количестве у новорожденных, чем у взрослых. После рождения, примерно до 1 −1,5 лет жизни, Le-генспецифическая трансфераза вырабатывается в большем количестве, чем Se-генспецифическая. На этом фоне синтезируются преимущественно иммунодоминантные Le a-сахара, фенотип Le(a +b −), и нередко иммунодоминантные Le b-сахара, фенотип Le(a +b + ) [62, 140].
Начиная с 2 −3-летнего возраста продукция трансфераз уравнивается в количественномотношении,ифенотипдетейLe(a +b −)иLe(a +b + )изменяетсянаLe(a −b + ).
MäkeläиMäkelä(154)показали,чтоплазмановорожденныхнетрансформирует эритроциты Le(a −b −) взрослых в Le(a + ), но эритроциты Le(a −b −) детей могут трансформироваться в Le(a + ) при инкубации их в плазме взрослых Le(a + ).
Низкий уровень трансферазной активности и, следовательно, низкая концентрация олигосахаридов Lewis в плазме новорожденных создают видимость