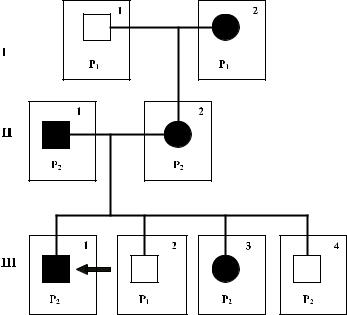
в этом случае ген Lu должен был иметься одновременно у 5 различных неродственных лиц, что представляется маловероятным. Характер наследования гена указы-
валнаегорецессивныйтип,ассоциированныйсХ-хромосомой.ВселицаLunull были мужчинами, среди представителей следующего поколения не было лиц указанного
фенотипа (см. рис. 8.4). Регуляторный локус был маркирован как XS, обычный часто встречающийся аллель – XS1, а редкий ингибиторный, вызывающий супрессию генов LU, – XS2. В данной семье мужчины с фенотипом Lunull были гемизиготными по ингибиторномуаллелю(XS2 / Y),женщины–гетерозиготными(XS1 / XS2).
Williamson и соавт. [148] привели случай аутоиммунной тромбоцитопении у мужчины, временно утратившего антигены Kell и имевшего антитела анти-Ku (KEL5). Его эритроциты нормально экспрессировали антигены Lu a, Lu b и антиген LW a системы LW. Через год экспрессия антигенов Kell нормализовалась, анти- Ku-антитела исчезли, однако имела место утрата антигенов Lutheran, а сыворотка крови содержала антитела анти-Lu3. Экспрессия антигена LW a резко снизилась.
Poole и соавт. [111] наблюдали еще одого больного группы Lu(a −b + ) с аутоиммунной тромбоцитопенической пурпурой. В течение заболевания его фенотип сменился на Lu(a −b −), экспрессия антигенов AnWj и LW оставалась нормальной. Сыворотка крови содержала анти-Lu-подобные антитела.
Другие антигены LU
Помимо антигенов Lu a, Lu b и Lu3 в систему Lutheran входят 16 других антигенов: 12 часто встречающихся, 2 редко встречающихся и 2 с частотой 50–80 % (см. табл. 8.1). Шесть из них являются продуктами аллельных генов и образуют антитетичные пары: Lu6 и Lu9, Lu8 и Lu14,Au a иAu b. Антигены Lu4, Lu5, Lu7, Lu12, Lu13, Lu20 и Lu21 отсутствуют на эритроцитах Lunull.
Антигены Lutheran локализуются в основном на гликопротеине Lu, многие слабо экспрессированы на эритроцитах новорожденных, не имеют рекомбина-
ций и кроссинговера (Daniels и соавт. [33, 38], Levene и соавт. [80], Parsons и соавт. [104], Reid и соавт. [116]).
Локализация детерминант Lu11, Lu16 и Lu17 на гликопротеине Lu не доказана, поэтому по отношению к ним используют обозначение «пара-Lutheran».
Антигены Lu11, Lu16 и Lu17 отсутствуют на эритроцитах Lunull и In(Lu) рецессивного типа. Антиген Lu17 присутствует на эритроцитах Lunull Х-ассоциированного типа.
БольшинствоантителLutheranневызываетгемолитическихосложненийиГБН.
Lu4
Bove и соавт. [12] нашли единственный образец антител анти-Lu4. Двое сибсов белой женщины группы Lu: −4, у которой были выявлены антитела, также имели фенотип Lu: −4. Все лица, кровь которых исследована с использованием указанной сыворотки(2700человек,преимущественнодоноров),имелифенотипLu4 +.
АнтигенLu4размещаетсянавторомIgSF-доменегликопротеинаLu(см.рис.8.1).