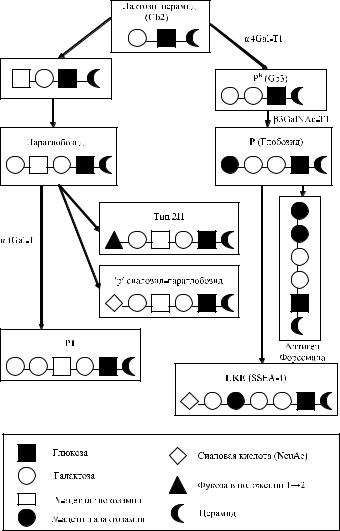
антигены Р и Р1 (Kannagi и соавт. [74]), что послужило основанием концепции «запретных антигенов», появляющихся в опухолях вопреки генотипу индивида, сфорсулированной Levine. По появлению этих антигенов можно судить о глубине трансформации нормальных клеток в процессе малигнизации. Levine высказал предположение, что антитела анти-РР1Р k, имевшиеся у больной, предотвращали дальнейший рост опухоли.
Необычные фенотипы
Race и Sanger [133] описали пациента, у которого присутствовали антитела антиРР1Р k. Эритроциты больного, Р1 −Р k −LKE −, реагировали с 9 сыворотками анти-Р1, полученнымиотлицР k,атакжес1из12сыворотоканти-РР1Р k инеагглютинирова- лисьсывороткойанти-Рбольногоэхинококкозом.Сывороткабольногореагировалас эритроцитамиР1,Р2 иР k,нобылаинертнавотношении16образцовэритроцитовр.
Allenисоавт.[3]обследовалибольногосослабымантигеномР(Р1 −Р k −),всыворотке которого присутствовали антитела анти-IP1 и анти-IP. Антиген I в эритроцитах этого человека также был необычным: компонент I D был ослаблен, I F – выражен нормально, экспрессия антигена i была повышена (см.Система Ii).
Issitt и соавт. [68] описали антитела анти-IP1, которые в серологических реакциях вели себя как анти-Р1 с той лишь разницей, что не реагировали с эритроцитами новорожденных Р1.
Booth [16], Ramos и соавт. [135] нашли двухфазные гемолизины со специфичностью анти-I ТP1.
Kundu и соавт. [88] нашли мужчину китайца с фенотипом Р2, эритроциты которого слабо реагировали с сыворотками анти-РР1Р k. В сыворотке крови антител не выявлено. Содержание глобозида, в том числе Gb3, в его эритроцитах составляло 30–40 % от нормального. Авторы высказали предположение, что возникновение подобного фенотипа обусловлено гомозиготностью индивида по дефектному аллелю Р k или эффектом другого гена, ингибирующего ген Р k.
В сыворотке крови 1 женщины были найдены агглютинины, которые давали выраженные реакции с эритроцитами Р1, Р2 и слабоположительные реакции с эритроцитами р (Shirey и соавт. [150]). Специфичность антител более напоминала анти-Р, чем анти-LKE, поскольку они в отличие от анти-LKE-антител ингибировались глобозидом.
Обследуемая имела фенотип Р1 + Р k + Р −. В то же время содержание глобозида в эритроцитах было снижено в 2, а Gb3 – в 4–6 раз по сравнению с нормой.
Не исключено, что антигены рассматриваемой системы, как и другие групповые антигены, могут быть парциальными.
Анти-р-антитела
До настоящего времени специфические анти-р-антитела не найдены и, возможно, не существуют в природе. Вместе с тем нечто близкое к анти-р- специфичности было обнаружено.