Материал: Билеты по физиологии+формулы+показатели организма
Таким способом моделируется невротическое состояние с преобладанием процесса возбуждения.
Перенапряжение и срыв подвижности основных корковых нервных процессов. Это достигается при:
- Переделке сигнального значения разных условных раздражителей (например, световой сигнал вместо ранее положительного подкрепления — получения пищи сопровождается последующим болевым воздействием).
- Ломке сложившегося динамического стереотипа (серии последовательных условных рефлексов).
Подобные воздействия обычно приводят к развитию невротических состояний с патологической подвижностью нервных процессов.
«Сшибка» рефлексов-инстинктов взаимоисключающего (противоположного) биологического значения. Осуществляется путём экстренного изменения качества подкрепляющего воздействия, например, подачей электрического тока в пол кормушки в момент пищевого подкрепления какого-либо сигнала либо воздействием сильного болевого раздражителя (биологически негативного) в момент полового акта.
Современные подходы к методам экспериментального воспроизведения неврозов у животных направлены на максимальное приближение к условиям их возникновения у человека. К таким методам относятся:
- Ограничение «рефлекса-инстинкта свободы» ( например, насильственная фиксация животного в станке).
- Нарушение естественного суточного режима питания или светоритма, связанных со сменой дня и ночи.
- Изменение привычных стадно-иерархических или стадно-половых отношений (например, у обезьян).
- Предварительнаяастенизация нервной системы (например, под влиянием хронического шума, ионизирующей радиации, изоляции животного от родителей в раннем детском возрасте).
Виды неврозов
- Невроз с патологической инертностью. Характеризуется частым развитием фобий. - Невроз с патологической лабильностью. Проявляется «суетливостью», незавершённостью действий, повышенной двигательной активностью. - Циркуляторный (циклический) невроз. Характеризуется хаотическим чередованием перечисленных выше разновидностей невроза.
Проявления экспериментальных неврозов.
- Расстройства ВНД. Они выражаются выпадением условных рефлексов, увеличением латентных периодов ответов на воздействия, трудностью или невозможностью выработки новых условных рефлексов и как следствие — адекватного приспособления к меняющимся условиям жизнедеятельности. Это ведёт к снижению адаптивных возможностей нервной системы и организма в целом, утрате индивидуальных черт реагирования, снижению «обучаемости» животных новым навыкам.
- Развитие так называемых фазовых состояний в нервной системе. Они характеризуются качественной и/или количественной неадекватностью peaгирования индивида на раздражители в зависимости от доминирующего в данный момент фазового состояния.
- Нарушение вегетативных функций при неврозах. Этот признак является постоянным, наиболее ранним и устойчивым проявлением неврозов. Изменение вегетативных функций, как правило,
Роль особенностей ВНД в возникновении неврозов.
Одни и те же экспериментальные воздействия нередко вызывают различные нарушения нервных процессов в высших отделах нервной системы. В большой мере это зависит от типа ВНД. В лаборатории И.П. Павлова была установлена зависимость вероятности возникновения и особенностей развития невроза от особенностей ВНД.
- Наиболее подвержен невротическим расстройствам меланхолик. Для него характерны быстрая истощаемость возбудительного процесса, слабость внутреннего коркового торможения, пассивность реакции на воздействие. Это предопределяет возникновение невроза в результате срыва основных корковых нервных процессов с развитием торможения и формированием пассивно-оборонительных реакций. - Холерик. Этот тип отличается сильным возбудительным процессом, слабым корковым торможением, активными реакциями на раздражители. Это обусловливает развитие невроза возбудительного типа с формированием активно-поисковых реакций. - Флегматик. Характеризуется развитием невроза с патологической подвижностью нервных процессов. - Сангвиник. Наиболее устойчив к воспроизведению неврозов в связи с высокой резистентностью его к различного рода патогенным агентам. Повышение силы раздражителя, «сшибка» инстинктов, увеличение деятельности и повторение воздействий могут привести к неврозу.
Заключение. Одной из наиболее распространённых форм патологии нервной системы являются неврозы. Невроз является реакцией личности на трудную, часто не разрешимую для него ситуацию. В целом в основе возникновения неврозов находится невротический конфликт, т.е. такое отношение личности к конкретной ситуации, которое делает невозможным и «непосильным» её рациональное решение. Термин «невроз» применяют для обозначения функциональных расстройств нервной системы — нарушений ВНД.
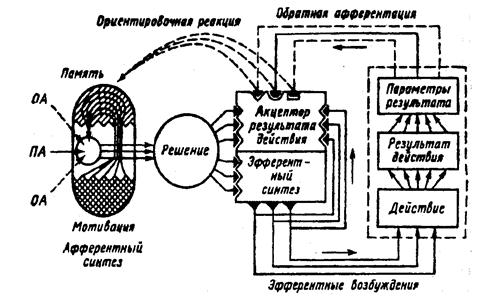
56. Учения Анохина о функциональных системах и саморегуляции функций. Структура целостности поведенческого акта с точки зрения теории Анохина.
От того, какой конечный результат нужен организму, в работу включаются в определенной последовательности определенные структуры мозга.
Функциональная система – это группа процессов и структур, объединенных для выполнения какой-либо функции организма. Ф.С. – это единица интегративной деятельности мозга. Например, функциональная система регуляции дыхания.
Функциональная система – динамическая, избирательно объединенная центрально-периферическая организация, деятельность которой направлена на достижение полезного для организма приспособительного результата.
Свойства, особенности функциональных систем:
1) динамична, способна к постоянной перестройке, к избирательному вовлечению мозговых структур для меняющихся поведенческих реакций;
2) структуры функциональных систем могут быть расположены как на большом, так и на близком расстоянии;
3) вовлечение структур в работу зависит от результата, которого надо достичь;
4) при нарушении функции в одной части системы, происходит срочное перераспределение активности во всей системе;
5) Ф.С. могут устранять, не принимать во внимание те афферентные воздействия, которые не имеют значения для организации конечной приспособительной реакции – принцип сужения афферентации, когда сохраняется только ведущая афферентация.
6) Если конечная цель не достигается, возможно включение дополнительной афферентации.
Функциональная система включает следующие элементы:
· Управляющее устройство – нервный центр;
· Выходные каналы нервного центра (нервы и гормоны);
· Исполнительные органы – эффекторы;
· Рецепторы результата (сенсорные рецепторы) – воспринимают информацию о параметрах процесса и его отклонение от оптимального уровня;
· Каналы обратной связи – с помощью импульсации от рецепторов или на основе изменения химического состава жидкостей информируют нервный центр об эффективности процесса. При этом корректируется деятельность всей функциональной системы.
· Пр: при увеличении давления раздражаются барорецепторы рефлексогенных сосудистых зон, увеличивается поток импульсов в центр кровообращения, изменяется эфферентная импульсация, что ведет к ослабления деятельности сердца и расширению сосудов. Если этого недостаточно включаются дополнительные механизмы – возрастает переход жидкости из кровеносного русла в межклеточную среду и т.д.
Различные константы внутренней среды организма взаимосвязаны – изменение одних констант может привести к изменению других констант. Пр: избыточное потребление воды сопровождается увеличением объема циркулирующей крови.
Все функциональные системы гомеостатического уровня объединяются в функциональную систему гомеостазиса.
Также для регуляции поведенческих реакций, при удовлетворении потребностей организма формируются функциональные системы. Их структуру также разработал Анохин и развивает Судаков К.В.
Структура функциональной системы поведенческого акта:
1. Афферентный синтез – процесс взаимодействия возбуждения, возникающего в головном мозге под влиянием обстановочной афферентации (сигналы об обстановке), с возбуждением, вызванным условным сигналом, в результате чего формируется действие.
2. Принятие решений – организм готовится к совершению действия, необходимого именно в данный момент, при этом исключаются остальные неподходящие варианты. Формируется программа действия.
3. Акцептор результата действия – включаются в работу эфферентные структуры, происходит осуществление самого действия. Обратная афферентация сигнализирует о результатах, которые сравниваются с запрограммированными в акцепторе действия. Если цель достигнута система перестает функционировать.
Схема центральной архитектоники поведенческого акта по Анохину П.К
57. Память и ее значение в формировании целостности приспособительных реакций. Особенности механизма кратковременной и долговременной памяти. Физиологические основы обучения.
Огромное значение для индивидуального поведения имеют обучение и память. Выделяют генотипическую или врождённую память ифенотипическую, т.е. приобретённую память. Генотипическая память является основой безусловных рефлексов и инстинктов. Фенотипическая память хранит информацию, поступающую в процессе индивидуальной жизни. Приобретённая память имеет 2 формы: чувственно-образную и логически-смысловую. Первая формируется в результате действия на анализаторы натуральных раздражителей (запах, вкус, цвет и т.д.), вторая - на основе абстрактных понятий (слова, формулы и т.д.). Чувственно-образную память делят по характеру раздражителей на зрительную, слуховую, вкусовую и т.д. Обе формы памяти постоянно взаимодействуют, образуя сложные ассоциации (например, название цветка ассоциируется с его внешним видом, запахом). Процесс запоминания происходит в четыре этапа: 1. Сенсорная память. В ней происходит кратковременное удержание сенсорной, т.е., поступившей в органы чувств информации. На этом этапе информация хранится доли секунды. В это время происходит анализ сигналов и большая часть информации переходит в кратковременную память, меньшая - сразу в промежуточную или долговременную. 2. Кратковременная память. Здесь информация находится до несколько минут. Ненужная информация отсюда удаляется, а имеющая значение переходит в промежуточную память. 3. Промежуточная память. В ней информация может храниться от нескольких десятков минут до нескольких лет. Неречевая информация из сенсорной памяти может сразу переходить в промежуточную память (инпринтинг - запечатлевание). Речевая же обязательно поступает через кратковременную память в промежуточную. Причём словесная информация закрепляется в ней лишь после нескольких повторений. 4.Долговременная память. В нее информация переходит из промежуточной, причём этот переход происходит во время быстрого сна. Первый этап запоминания, т.е. сенсорная память является результатом возникновения нервных импульсов в периферических рецепторах, их распространения по проводящим путям в корковый отдел анализатора и процессов высшего синтеза в коре. Кратковременная память обусловлена поступлением нервных импульсов в гиппокамп, где выделяется главная и отбрасывается ненужная информация. После этого информация поступает в замкнутые нейронные сети, где происходит циркуляция или реверберация нервных импульсов. Переход информации в промежуточную и долговременную память происходит в коре полушарий на основе более тонких механизмов. Следы памяти в нейронных цепях коры формируются в результате 2-х процессов: 1. 3а счёт усиления или потенциации нервных импульсов в межнейронных синапсах. Потенциация происходит в результате увеличения количества выделяемого нейромедиатора и числа постсинаптических рецепторов. 2. Благодаря структурным изменениям мембран и органелл нейронов. Эти изменения синаптической передачи и мембран являются следствием предшествующей реверберации. Данными процессами обеспечивается промежуточная и долговременная память. Кроме того предложены другие теории долговременной памяти. 1. Химическая теория. В её основе лежат опыты с "транспортом памяти" (обучение животных - введение экстракта их мозга необученным животным, опыты со скотофобином). Согласно этой теории информация хранится в специальных белках, синтезируемых нейронами. 2. Теория хранения энграммы в ДНК. Предполагают, что ДНК программирует необходимые изменения структуры и свойств синапсов и таким образом обеспечивает перестройку нейронных цепей в процессе запоминания. Нарушения памяти; 1. Ретроградная амнезия - утрата способности мозга к извлечению информации, поступившей в мозг до момента экстремального воздействия на него, потеря информации, накопленной до травмы головного мозга или сильного опьянения. Под гипнозом эту информацию можно извлечь. 2. Антеретроградная амнезия - неспособность к запоминанию новой информации. В клинике синдром Корсакова. Память на отдалённые события сохранена, а недавние быстро забываются. Хр. Алкоголизм. Поражение гипнокампа
Каковы же физиологические механизмы, лежащие в основе «кратковременной» и «долговременной» памяти?
Еще в тридцатые и сороковые годы было сделано наблюдение, которое дало основу для того, чтобы высказать гипотезу о природе тех нервных процессов, которые лежат в основе «кратковременной» памяти.
Морфологическими и морфофизиологическими исследованиями американских нейрофизиологов Лоренте-де-Но и Мак Кэллока было установлено, что в коре головного мозга существуют аппараты, позволяющие возбуждению длительно циркулировать по замкнутым цепям. Основой служил тот факт, что у аксонов отдельных нейронов существуют веточки, которые возвращаются к телу этого же нейрона и либо непосредственно соприкасаются с ним, либо соприкасаются с отдельными дендритами этого же нейрона; этим самым создается основа для постоянной циркуляции возбуждений в пределах замкнутых круговых цепей, илиреверберирующих кругов возбуждения. Этим простейшим механизмом, однако, дело не ограничивается. Есть все основания думать, что в нервной системе существуют и более сложные аппараты «нейронных сетей», осуществляющих устойчивые реверберационные круги возбуждения. Такими аппаратами являются функциональные комплексы нейронов, соединенные друг с другом «вставочными» нейронами, или нейронами с короткими аксонами, функция которых, по-видимому, состоит в том, чтобы передавать возбуждение от одного нейрона к другому, обеспечивая длительное протекание возбуждения по более сложным сетям, или «реверберационным кругам».
Некоторые исследователи считают, что «реверберационные круги» возбуждения и являются нейрофизиологической основой «кратковременной» памяти. Существенным механизмом сохранения следов оказывается, по этим предположениям, механизм синаптической передачи возбуждения, который и обеспечивает переход возбуждения с одного нейрона на другой и дает возможность осуществлять длительное сохранение возбуждения, протекающего по «реверберациоиным кругам».
Согласно этой теории шок разрушает протекание возбуждения по реверберационным кругам и приводит к исчезновению тех следов, которое сохранялись благодаря этому протеканию возбуждения.
Процесс циркуляции возбуждения по «реверберационным кругам» не является, однако, единственно возможным механизмом сохранения следов. Факты, полученные многими исследователями, заставили предположить, что механизм сохранения следов теми глубокими биохимическими изменениями, которые могут ходить не только в синапсах (места передачи возбуждения от одних нейронов к другим), но и в самих телах нейронов и их отдельных органах (ядрах, метахондриях).
Еще в 1959 г. шведский исследователь Хиден показал, что каждое раздражение нервных клеток приводит к заметному повышению содержания рибонуклеиновой кислоты (РНК), в то время как длительное отсутствие раздражений уменьшает содержание РНК. Дальнейшие наблюдения Хидена и его сотрудников заставили высказать предположение, что изменения РНК носят специфический характер и могут предположительно рассматриваться как биохимический механизм сохранения следов памяти. Основой для предположения является тот факт, что изменения РНК, вызванные определенными воздействиями, могут быть очень специфичны, и что различные воздействия могут вызывать разные модификации РНК.
Было высказано предположение, что число возможных изменений молекул РНК под влиянием различных воздействий измеряется огромным числом – 1015-1020, и, таким образом, РНК оказывается в состоянии сохранять огромное число различных кодов. Как предполагали эти исследователи, повторное появление этого раздражителя приводит к тому, что специфически измененная РНК начинает «резонировать» именно этому раздражению, а способность специфического резонирования именно данному раздражению и является основой того, что нервная клетка, сохраняющая след полученного воздействия, начинает «узнавать» это воздействие, отличая его от всякого другого.
Такое специфическое изменение РНК под влиянием различных воздействий и дало основание исследователям для предположения, что оно является биохимической основой памяти.
Предположение об участии РНК в сохранении следов памяти было подтверждено рядом наблюдений. К ним относятся наблюдения, проведенные известным американским физиологомМореллем, которой показал, что повышение содержания РНК, вызываемое повторным раздражением определенного участка мозга, проявляется не только в этом очаге, но и в симметричном ему пункте другого полушария. Это означает не только то, что реверберационные круги возбуждения могут охватывать очень большие зоны мозга, распространяясь и на противоположное полушарие, но и то, что в этом симметричном «зеркальном фокусе», который не испытывал никакого прямого влияния раздражителя, возникает повышенное содержание РНК, очевидно, указывающее на создавшуюся в нем готовность к повторным возбуждениям.
К числу их относятся наблюдения, сделанные при помощи электронной микроскопии, показавшие, что по мере формирования следов навыка в соответствующих нейронах животного можно наблюдать увеличение числа мельчайших везикул (пузырьков), содержащих повышенную концентрацию ацетилхолина, способствующего перенесению импульса в синапсах, в то время как длительное отсутствие раздражений уменьшает их количество.
К числу таких наблюдений относятся и факты, которые показали, что следы информации, усвоенной животным, могут быть переданы другому животному гуморальным путем через посредство измененной РНК, и наоборот, разрушение РНК (ее растворение рибонуклеазой) приводит к разрушению этих следов.
Эти наблюдения вызвали оживленную дискуссию, и мы приводим краткие данные, отмечая, что их проверка и окончательная оценка еще являются делом будущего.
Данные о возможном участии РНК как в хранении, так и в передаче информации, были впервые получены американским исследователем Мак Коннеллом. Этот исследователь вырабатывал у плоских червей (планарий) навык избегать света. Такое обучение требовало значительного числа проб. После этого планария разрезалась на две части, каждая из которых постепенно регенерировала, превращаясь в целое животное. Когда регенерировавшие особи снова начинали обучаться той же процедуре, оказывалось, что обучение как регенерировавшего головного, так и регенерировавшего хвостового шипа требует втрое меньшего количества тренировочных проб. Следовательно, сохранение следа памяти происходит не за счет оставшихся нейронов переднего ганглия (который у хвостового конца заново регенерировал), а за счет гуморальных (биохимических) сдвигов, сохранившихся во всех тканях тела. Характерно, что, если оба конца планарии, у которой был выработан соответствующий навык, опускались в раствор рибонуклеазы, разрушавшей РНК, следы полученного навыка исчезали, и регенерировавшие черви требовали для повторного обучения такого же количества новых тренировочных опытов, как и необученные особи.
Эти опыты, по мнению авторов, подтверждают участие РНК в сохранении следов памяти.
Дальнейшие опыты, проведенные Мак Конпелом и другими исследователями, создали впечатление, что измененная РНК может не только сохранять следы полученной информации, но и передавать их гуморальным путем другим особям. Для того, чтобы показать это, Мак Коннел сначала вырабатывал соответствующий навык у группы планарии, а потом скармливал экстракт из тел обученных планарии необученным планариям. По данным, которые приводит исследователь, в результате этого опыта необученные планарии начинали значительно быстрее вырабатывать тот специфический навык, который ранее вырабатывался у обученных планарии и, по-водимому, передавался им гуморальным путем посредством специфически измененной РНК, хранящей следы выработанной модификации поведения.
Подобные опыты были проведены на ряде животных (в том числе на крысах, в мозг которых вводился экстракт из размельченного мозга ранее обучавшихся крыс), и авторы, проводившие эти опыты, высказали предположение, что и в этих случаях РНК участвует не только в сохранении следов от полученной информации, но в передаче этой информации другим особям гуморальным (биохимическим) путем.
Как уже было сказано выше, эти опыты вызвали горячую дискуссию, и еще трудно утверждать, что их результаты подтвердятся дальнейшими исследованиями. Возникает существенный вопрос: ограничивается ли изменение РНК, возникающее в результате раздражения, одними лишь нейронами, или же в процесс сохранения следов вовлекаются и другие ткани мозга? Этот вопрос привлек внимание исследователей.
Как известно, в состав ядер подкорковых образований, как и в состав коры, кроме нейронов входит еще и глия, которая облегает нервные клетки плотной губчатой массой. В течение длительного времени глия считалась лишь опорной тканью мозга, однако за последнее время стало ясно, что она имеет и другие, гораздо более сложные функции, участвуя как в обменных процессах, так и в регуляции процессов возбуждения, протекающего в нервных аппаратах, а возможно, и в процессе сохранения следов тех возбуждений, которые возникают в нервной ткани мозга. Известно также, что число глиальных клеток в 10 раз больше, чем число нервных клеток; в отличие от нервных клеток, которые не делятся при жизни, глиальные клетки продолжают делиться и число их увеличивается в онтогенезе. Характерно, что по мере развития существенно возрастает отношение массы нервных клеток ко всей массе серого вещества, к которой относятся и глиальные клетки.
Глиальные клетки плотно облегают нервные клетки, и, по выражению Хидсна, «занимают стратегическое положение между нервными клетками и кровеносными капиллярами». Электрические потенциалы возникают в них во много сотен раз медленнее, чем в нервных клетках, а биохимические изменения, происходящие в них под влиянием раздражений, находятся в обратных отношениях к биохимическим изменениям, происходящим в нервных клетках: в начале раздражения в нервных клетках (нейронах) количество РНК увеличивается, а в окружающей глии – уменьшается, и наоборот, по окончании действия раздражителя количество РНК в нервной клетке быстро падает, а в клетках окружающей глии – возрастает. Поэтому возникновение медленных потенциалов, которым нейрофизиология придает особенно большое значение, связывается сейчас не только с работой нейронов, но и с работой глии.
Все это заставляет предполагать, что глия придает стабильность процессам, возникающим в нервной клетке, оказывает модулирующее влияние на протекание возбуждений и, возможно, непосредственно участвует в хранении следов тех возбуждений, которые возникают в нейронах.
Циркуляции возбуждений по реверберационным кругам и указаний на биохимические сдвиги, возникающие под влиянием раздражений, доходящих до нервной ткани, все же недостаточно для объяснения механизмов, лежащих в основе долговременной памяти. Поэтому некоторые исследователи считают необходимым искать механизмы долговременной памяти в некоторых морфологических изменениях, возникающих в синаптическом аппарате нейронов, и высказывают предположение, что именно эти морфологические новообразования являются субстратом долговременной памяти. Еще раньше известный морфофизиолог А. Капперс указал, что рост аксонов и денд-ритов не случаен и отростки нейрона ориентируются в направлении протекающего возбуждения. Это явление, которое А. Капперс назвал «нейробиотическим», подтвердилось при дальнейших наблюдениях. Сейчас ученые полагают, что направление роста отростков нейронов в значительной мере определяется их функционированием и теми «программами», которые зависят от кода возбуждения и лежат в основе их деятельности.