Материал: Билеты по физиологии+формулы+показатели организма
Ядра таламуса функционально по характеру входящих и выходящих из них путей делятся на специфические, неспецифические и ассоциативные.
К специфическим ядрам относятся переднее вентральное, медиальное, вентролатеральиое, постлатеральное, постмедиальное, латеральное и медиальное коленчатые тела. Последние относятся к подкорковым центрам зрения и слуха соответственно.
Основной функциональной единицей специфических таламических ядер являются «релейные» нейроны, у которых мало дендритов и длинный аксон; их функция заключается в переключении информации, идущей в кору большого мозга от кожных, мышечных и других рецепторов.
От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки III—IV слоев коры большого мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности, так как ядра таламуса, как и кора большого мозга, имеют соматотопическую локализацию. Отдельные нейроны специфических ядер таламуса возбуждаются рецепторами только своего типа. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют сигналы от интерорецепторов зон проекции блуждающего и чревного нервов, гипоталамуса.
|
Латеральное коленчатое тело имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
В медиальное коленчатое тело (МТК) поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферетные пути от медиальных коленчатых тел идут в височную зону коры большого мозга, достигая там первичной слуховой области коры. МКТ имеет четкую тонотопичность. Следовательно, уже на уровне таламуса обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Ассоциативные ядра таламуса представлены передним медиодорсальным, латеральным дорсальным ядрами и подушкой. Переднее ядро связано с лимбической корой (поясной извилиной), медиодорсальное — с лобной долей коры, латеральное дорсальное — с теменной, подушка — с ассоциативными зонами теменной и височной долями коры большого мозга.
Основными клеточными структурами этих ядер являются мультиполярные, биполярные трехотростчатые нейроны, т. е. нейроны, способные выполнять полисенсорные функции. Ряд нейронов изменяет активность только при одновременном комплексном раздражении. На полисенсорных нейронах происходит конвергенция возбуждений разных модальностей, формируется интегрированный сигнал, который затем передается в ассоциативную кору мозга. Нейроны подушки связаны главным образом с ассоциативными зонами теменной и височной долей коры большого мозга, нейроны латерального ядра — с теменной, нейроны медиального ядра — с лобной долей коры большого мозга.
Неспецифические ядра таламуса представлены срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексами, ретикулярным ядром, перивентрикулярной и центральной серой массой. Нейроны этих ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из РФ ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса.
Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния. Нарушение функции неспецифических ядер затрудняет появление веретенообразной активности, т. е. развитие сонного состояния.
Сложное строение таламуса, наличие в нем взаимосвязанных специфических, неспецифических и ассоциативных ядер, позволяет ему организовывать такие двигательные реакции, как сосание, жевание, глотание, смех. Двигательные реакции интегрируются в таламусе с вегетативными процессами, обеспечивающими эти движения.
Конвергенцию сенсорных стимулов в таламус обусловливает возникновение так называемых таламических неукротимых болей, которые возникают при патологических процессах в самом таламусе.
Таламус участвует в передаче и переработке информации: специфические ядра таламуса обеспечивают анализ локализации болевого раздражения, его силы и длительности, неспецифические ядра таламуса участвуют в формировании мотивационно-аффективного аспекта боли.
49. Гипоталамус. Классификация и характеристика ядерных групп. Участие гипоталамуса в регуляции вегетативных функций и формировании эмоций и мотиваций.
Серое в-во гипоталамической области группируется в ядра (около 30), носящие названия, в соответствии с их анатомическим положением : «Серобугорные ядра», «ядро воронки», « медиальное» и «латеральное» - ядра сосцевидных тел, а так же в соответствии с расположением их в этой области: «Заднегипотиламич» - самое выраженное и массивное, «медиальные» ядра; «супраоптическое» и «паравентикулярное», располагающиеся в передней части области и тесно связанные с гипофизом.
Нерв кл, составляющие серое вещество неоднородны.Часть нейронов настолько изменена, что они не столько биоэлектрически активны, сколько специализировались на секреции (синтез и выделение в кровь) нейрогормонов, регулирующих активность всех желез внутренней секреции организма. Основное их количество находится в передних ядрах, связанных с гипофизом. Другая часть – изменена таким образом, что из раздражимость связана не с биоэлектропотенциалами соседних нейронов,а с изменением отдельных химических параметров внутренней среды.Их дендриты связаны с кровеносным руслом, и рецептируют температуру и состав крови (кол-во глюкозы,мин.солей,воды,углекислоты).Эти клетки входят в состав ядер – центров, регулирующих отдельные виды обмена веществ (углнводного,водно-солевого) и называются интрорецепторами.
Белое вещество области представлено в основном тонкими прослойками между ядрами,а так же располагается по наружн поверхности обеспечивая как внутренние взаимодействия так и связь области с окруж структурами.
В функциональном плане эта область является центральной структурой, обеспечивающей (А)«стратегическую» организацию вегетативных процессов в соответствии с внутренними потребностями организма, и (Б) оптимизирует работу внутренних органов и обменные процессы в связи с постоянно изменяющимися условиями внешней среды, т.е. с внешними возможностями. За счет тесных связей с ретикулярной активирующей системой ствола мозга гипоталамические центры определяют (В) ритм активности практически всех процессов жизнедеятельности – от суточного, до сезонного. За счет наличия здесь видоизмененных нейронов, секретирующих нейрогормоны, и прямых связей с гипофизом и шишковидной железой, гипоталамус обеспечивает (Г) взаимосодействие нервной и гуморальной систем в регуляции функций организма.
Субъективно, активность ядер - гипоталамических центров ощущается как возникновение той или иной внутренней потребности - мотивации – (пищевой, оборонительно-исследовательской, половой), направляющей текущее поведение. При удовлетворении возникшей потребности в случае благоприятных внешних условий, активность центров перестраивается, что субъективно переживается как положительная эмоция (радость, удовольствие и пр.). Невозможность удовлетворить ту или иную потребность, т.е. обеспечить регуляцию обменных процессов в создавшихся условиях внешней среды, активизирует соответствующий центр, который начинает оказывать доминирующие влияния на остальные мозговые структуры. Это определяет перестройку текущего поведения, изменяет внутренне состояние, вплоть до состояния стресса, а субъективно ощущается как отрицательная эмоция.
Ядерные структуры гипоталамуса группируются в функциональные центры в соответствии с тем, какие виды обменных процессов они контролируют, причем каждый центр состоит из двух отделов – один ускоряет, а другой замедляет их протекание, т.е. являющихся антагонистами, а морфологически они располагаются в противоположных частях этого образования.
Выделяют центры голода/насыщения, жажды/водного насыщения, термопродукции\термоотдачи, центры полового возбуждения\пассивного состояния, центры ориентировочно-исследовательского поведения: агрессии/ затаивания и пр., которые направляют поведение на достижение соответствующих результатов, причем считается, что центры возбуждающего характера, например, центр голода, располагаются в латеральных отделах, а насыщения – в медиальных, хотя точная функциональная локализация не всегда может быть установлена. Ядра серобугорной области и сосцевидных тел участвуют в организации полового поведения и обеспечивают баланс температуры тела. Также в экспериментах на животных были выявлены центры «удовольствия» не связанные с какой либо конкретной мотивацией, возбуждение которых вызывает эйфорию.
Нервные центры гипоталамуса (ядра серого вещества) непосредственно связаны с нижележащими центрами продолговатого мозга и спинного мозга, обеспечивающими вегетативную регуляцию отдельных органов (дыхательный центр, сосудодвигательный и пр.) При этом имеется определенное морфологическое соответствие. Передняя группа ядер образует «трофотропную» систему, регулирующую обменные процессы с целью накопления и восстановления энергетических ресурсов, и связана с парасимпатическим отделом вегетативной нервной системы. Эта система активируется в спокойном состоянии, например, во сне, и эффекты ее возбуждения проявляются в активизации органов пищеварительной системы и угнетении практически всех остальных: дыхание становится редким и глубоким, мышцы расслабляются, снижается артериальное давление и пр.. Задняя группа ядер образует «эрготропную» систему, обеспечивающую регуляцию обменных процессов и работу внутренних органов в условиях постоянно изменяющейся внешней среды, в стрессовых ситуациях. Она связана с симпатическим отделом, и эффекты ее возбуждения противоположны, и также она активирует систему термоотдачи и водносолевого обмена.
50. Лимбическая система. Структурно-функциональная организация. Ее афферентные и эфферентные связи. Роль лимбической системы в формировании биологический мотиваций и эмоций.
Структурно-функциональная организация. В лимбическую систему включают образования древней коры (обонятельная луковица и бугорок, периамигдалярная и препериформная кора), старой коры (гиппокамп, зубчатая и поясная извилины), подкорковые ядра (миндалина, ядра перегородки), и этот комплекс рассматривается по отношению к гипоталамусу и ретикулярной формации ствола как более высокий уровень интеграции вегетативных функций. Кроме вышеназванных структур в лимбическую систему в настоящее время также включают гипоталамус, ретикулярную формацию среднего мозга. Афферентные входы в лимбическую систему осуществляются от различных областей головного мозга, а также через гипоталамус от ретикулярной формации ствола, которая считается главным источником ее возбуждения. В лимбическую систему поступают импульсы от обонятельных рецепторов по волокнам обонятельного нерва - коркового отдела обонятельного анализатора. Эфферентные выходы из лимбической системы осуществляются через гипоталамус на нижележащие вегетативные и соматические центры ствола мозга и спинного мозга. Лимбическая система оказывает восходящие возбуждающие влияния на новую кору (преимущественно ассоциативную).
Структурной особенностью лимбической системы является наличие хорошо выраженных кольцевых нейронных цепей, объединяющих различные ее структуры. Эти цепи дают возможность длительной циркуляции (реверберации) возбуждения, что является механизмом его пролонгирования, повышения проводимости синапсов и формирования памяти. Реверберация возбуждения создает условия для сохранения единого функционального состояния структур замкнутого круга и навязывает это состояние другим структурам мозга.
Функции. Лимбическая система после получения информации о внешней и внутренней среде организма, сравнения и обработки этой информации запускает через эфферентные выходы вегетативные, соматические и поведенческие реакции, обеспечивающие приспособление организма к внешней среде и сохранение внутренней среды на определенном уровне. В этом состоит наиболее общая функция лимбической системы. Основными частными функциями лимбической системы являются следующие.
1. Регуляция висцеральных функций. В этой связи лимбическую систему иногда называют висцеральным мозгом. Данная функция осуществляется преимущественно посредством гипоталамуса, который является диэнцефалическим звеном лимбической системы. О тесных эфферентных связях лимбической системы с внутренними органами свидетельствуют разнообразные разнонаправленные изменения их функций при раздражении лимбических структур, особенно миндалин: происходит повышение или понижение частоты сердечных сокращений, усиление и угнетение моторики и секреции желудка и кишечника, секреции гормонов аденогипофизом. 2. Формирование эмоций. Эмоции являются субъективным компонентом мотиваций - состояний, запускающих и реализующих поведение, направленное на удовлетворение возникших потребностей. Через механизм эмоций лимбическая система улучшает приспособление организма к изменяющимся условиям среды. Ключевой структурой для возникновения эмоций является гипоталамус. Выделяют собственно эмоциональные переживания и его вегетативные и соматические проявления. Эти компоненты эмоций могут иметь относительную самостоятельность в проявлениях: выраженные субъективные переживания могут сопровождаться небольшими соматическими и вегетативными проявлениями. Напротив, при псевдоэмоциях яркие внешние проявления в виде двигательных мимических реакций могут протекать без существенных субъективных переживаний. Гипоталамус при этом представляется структурой, ответственной преимущественно за вегетативные проявления эмоций - изменение частоты и силы сердечных сокращений, артериального давления, дыхания. Важную роль в возникновении эмоций играют поясная извилина и миндалина. Электрическая стимуляция миндалины у человека вызывает преимущественно отрицательные эмоции - страх, гнев, ярость. Напротив, двустороннее удаление миндалин в эксперименте на обезьянах резко снижает их агрессивность, повышает тревожность, неуверенность в себе. У таких животных нарушается способность оценивать информацию (особенно зрительную и слуховую), поступающую из окружающей среды, и связывать эту информацию со своим эмоциональным состоянием. В результате нарушается нормальное взаимодействие организма с окружающей средой, в том числе зоосоциальные отношения с другими особями в группе. Поясная извилина выполняет роль интегратора различных систем мозга, участвующих в формировании эмоций. 3. Лимбическая система участвует в процессах памяти и обучения. Особо важную роль играют гиппокамп и связанные с ним задние зоны лобной коры. Их деятельность необходима для консолидации памяти - перехода кратковременной памяти в долговременную. Электрофизиологической особенностью гиппокампа является его уникальная способность отвечать на стимуляцию длительной (в течение часов, дней и даже недель) посттетанической потенциацией, которая приводит к облегчению синаптической передачи и служит основой формирования памяти. Ультраструктурным признаком участия гиппокампа в образовании памяти является увеличение числа шипиков на дендритах его пирамидных нейронов в период активного обучения, что свидетельствует об усилении синаптической передачи информации, поступающей в гиппокамп.
Принципы организации поведенческих реакций (системная архитектоника поведенческого акта)
Вместе с тем, потребность и мотивация, сами по себе не способны сформировать полноценный поведенческий акт. В этом плане характерно высказывание И.М. Сеченова - "Голод, - писал И.М. Сеченов, - способен поднять животное на ноги, способен придать поискам более или менее страстный характер, но в нем нет никаких элементов, чтобы направить движение в ту или иную сторону и видоизменять его сообразно требованиям местности и случайных встреч" . Только после достижения того или иного адаптивного результата доминирующая мотивация оказывается способной активировать системные механизмы поведения и сформировать функциональную систему, определяющую тот или иной целенаправленный поведенческий акт.
Такие же закономерности проявляются при освоении человеком любых новых навыков и в решении новых задач.
Процессы подкрепления проявляют свою системообразующую роль уже в пренатальном онтогенезе, когда для восприятия адаптивных результатов в соответствующих тканях плода на генетически детерминированной основе опережающе созревают специальные рецепторы. На эти рецепторы начинают действовать информационные молекулы подкрепления, циркулирующие в крови матери. Под влиянием этих молекул уже в утробе материнского организма у плода складываются функциональные системы гомеостатического уровня.
Другая группа рецепторов опережающе созревает по отношению к полезным приспособительным результатам поведения, которые на ранних этапах постнатального развития играют решающую роль для пост-натального выживания новорожденных. В этом случае после рождения формирование функциональных систем происходит на основе механизма запечатления – импринтинга.
Изложенное свидетельствует о том, что полезные приспособительные результаты выступают в роли систем ообразующих фактороз становления функциональных систем разного уровня организации как при их созревании в онтогенезе, так и в процессах индивидуального обучения.
Обратим внимание на одно важное обстоятельство. Под влиянием подкрепления в центральной нервной системе живых существ формируется аппарат предвидения свойств потребного результата и способов его достижения - акцептор результата действия. Вследствие этого при возникновении той или иной потребности субъекты начинают предвидеть свойства потребного результата. Формируются специальные программы поведения. Поведение приобретает опережающий действительные собыгия характер и становится направленным на будущие потребные результаты.
Как правило, подкрепление развертывается во временной динамике. Временная динамика подкрепления складывается в процессе нередко продолжительного достижения субъектами, испытывающими ту или иную потребность, часто многочисленных промежуточных и только после этого - конечного результатов, удовлетворяющих эту потребность. Одни результаты поведения при этом могут приближать субъекта к удовлетворению исходной потребности, иногда в какой-то степени удовлетворять ее. Другие результаты - наоборот, препятствуют удовлетворению субъектом доминирующей потребности. При взаимодействии субъекта с разнообразными промежуточными и конечным результатами при обучении происходит формирование и ооогащение аппарата акцептора результата действия путем построения широко разветвленных по структурам мозга динамических энграмм подкрепления.
51. Базальные ганглии, их афферентные и эфферентные связи. Роль базальных ядер в формировании мышечного тонуса и сложных двигательных актов. Последствия повреждения базальных ганглий и их связей.
Базальные ганглии, названные так гистологами прошлого века, представляют собой структуры ядерного типа, которые располагаются в толще белого вещества переднего мозга ближе к его основанию.
У млекопитающих к базальным ганглиям относятся сильно вытянутое в длину и изогнутое хвостатое ядро и заложенное в толще белого веществачечевицеобразное ядро. Двумя белыми пластинками оно подразделяется на три части: наиболее крупную, лежащую латерально скорлупу ибледный шар, состоящий из внутреннего и внешнего отделов.
Эти анатомические образования формируют так называемуюстриопаллидарную систему, которая по филогенетическим и функциональным критериям разделяется на древний палеостриатум и неостриатум. Палеостриатум представлен бледным шаром, анеостриатум, появляющийся впервые у рептилий, состоит из хвостатого ядра и скорлупы, которые объединяются под названием полосатого тела или стриатума. Хвостатое ядро и скорлупа связаны анатомически и характеризуются чередованием белого и серого вещества, что оправдывает возникновение термина полосатое тело.
К стриопаллидарной системе часто относят также субталамическое ядро (люисово тело) и черную субстанцию среднего мозга, которые образуют с базальными ганглиями функциональное единство. Полосатое тело состоит главным образом из мелких клеток, аксоны которых направляются к бледному шару и черной субстанция среднего мозга.
Полосатое тело является своеобразным коллектором афферентных входов, идущих к базальным ганглиям. Главными источниками этих входов служат новая кора (преимущественно сенсомоторная), неспецифические ядра таламуса и дофаминэргические пути от черной субстанции.
В противоположность полосатому телу бледный шар состоит из крупных нейронов и является сосредоточением выходных, эфферентных путей стриопаллидарной системы. Аксоны локализованных в бледном шаре нейронов подходят к различным ядрам промежуточного и среднего мозга, в том числе и к красному ядру, где начинается
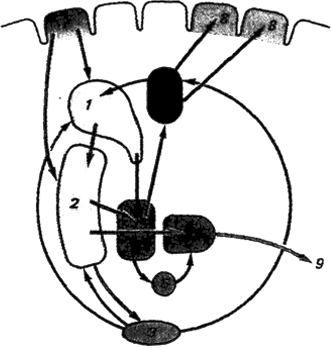 Рис.
3.15. Афферентные и эфферентные связи
(шальных ганглиев: 1 - хвостатое ядро, 2
- скорлупа, 3 - черная субстанция, 4 -
таламус, 5, 5' - внутренний и внешний отделы
бледного шара, б - субталамическое ядро,
7 - сенсомоторная кора, 8 - двигательная
кора, 9 - пути к стволовой части мозга;
синими стрелками обозначены афферентные
пути, красными - эфферентные, черными -
межъядерные взаимодействия
Рис.
3.15. Афферентные и эфферентные связи
(шальных ганглиев: 1 - хвостатое ядро, 2
- скорлупа, 3 - черная субстанция, 4 -
таламус, 5, 5' - внутренний и внешний отделы
бледного шара, б - субталамическое ядро,
7 - сенсомоторная кора, 8 - двигательная
кора, 9 - пути к стволовой части мозга;
синими стрелками обозначены афферентные
пути, красными - эфферентные, черными -
межъядерные взаимодействия
руброспинальный тракт экстрапирамидной системы двигательной регуляции.
Другой важный эфферентный тракт идет от внутреннего отдела бледного шара к передневентральному и вентролатеральному ядрам таламуса, а оттуда продолжается к двигательным областям коры головного мозга. Наличие этого пути обусловливает многозвенную петлеобразную связь между сенсомоторными и двигательными областями коры, которая осуществляется через полосатое тело и бледный шар к таламусу. Примечательно, что в составе этого стриапаллидоталамокортикального пути базальные ганглии выполняют роль афферентного звена по отношению к моторным областям коры головного мозга. Многочисленные связи стриопаллидарной системы с различными отделами мозга свидетельствуют об ее участии в процессах интеграции, однако до настоящего времени в знании о функциях базальных ганглиев остается много невыясненного.
Базальные ганглии играют важную роль в регуляции движений и сенсомоторной координации. Известно, что при повреждении полосатого тела наблюдается атетоз - медленные червеобразные движения кистей и пальцев рук. Дегенерация клеток стриатума вызывает также другое заболевание - хорею, выражающуюся в судорожных подергиваниях мимических мышц и мускулатуры конечностей, которые наблюдаются в покое и при выполнении произвольных движений. Однако попытки выяснить этиологию этих нарушений в экспериментах на животных не дали результатов. Разрушение хвостатого ядра у собак и кошек не приводило к возникновению гиперкинезов, характерных для описанных выше заболеваний.
Локальное электрическое раздражение некоторых участков стриатума вызывает у животных так называемые циркуляторные двигательные реакции, характеризующиеся поворотом головы и
туловища в сторону, противоположную раздражению. Раздражение других участков полосатого тела, напротив, приводит к торможению двигательных реакций, вызванных различными сенсорными раздражениями.
Наличие определенных расхождений между данными эксперимента и клиники, по-видимому, свидетельствует о возникновении системных нарушений механизмов регуляции движений при патологических процессах в базальных ганглиях. Очевидно, эти нарушения связаны с изменениями функции не только полосатого тела, но и других структур. В качестве примера можно рассмотреть возможный патофизиологический механизм возникновения дрожательного паралича - синдрома Паркинсона. Этот синдром связан с повреждением базальных ганглиев и характеризуется комплексом таких симптомов, как акинезия - малая подвижность и затруднения при переходе от покоя к движению; восковидная ригидность, или гипертонус, не зависящий от положения суставов и фазы движения; статический тремор, наиболее выраженный в дистальных отделах конечностей.
Все эти симптомы, согласно современным представлениям, обусловлены гиперактивностью базальных ганглиев, которая возникает при повреждении дофаминэргического (по всей вероятности, тормозного) пути, который идет от черной субстанции к полосатому телу. Таким образом, этиология синдрома Паркинсона обусловлена дисфункцией как полосатого тела, так и структур среднего мозга, которые функционально объединены в стриопаллидарную систему.