Фосфорилування-дефосфорилування гістонів
здійснюється в цитоплазмі та ядрі (гідроксильні групи серину і треоніну гістонів Н1, Н3).
Негістонові білки підлягають ковалентній модифікації значно більшою мірою, ніж гістони.
Ацетильні, фосфорні, метильні, глікозильні, АДФ-рибозильні радикали з’єднуються з бокови-
ми ланцюгами амінокислотних залишків у поліпептидних ланцюгах. Протягом різних фаз клітинного циклу закономірності ковалентної модифікації гістонів і негістонових білків можуть бути різними.
Синтез ДНК і S-фаза клітинного циклу
У тваринних клітинах, у тому числі в клітинах людини, реплікація ДНК-геному здійснюється у певний період клітинного (мітотичного) цик-
лу, який називають синтетичним, або S-фазою.
Зазвичай вона відокремлена від мітотичної фази несинтетичними періодами G1 (від англ. gap — проміжок) і G2, які знаходяться до та після S-фази. У тваринних клітин інтервал між мітозами (клітинний цикл) дорівнює приблизно 24 год. За цей час клітина проходить чотири фази життєвого циклу:
—G1-фазу початкового росту, де відбувається синтез мРНК, білків та інших компонентів клітини. У деяких клітин у клітинному циклі
може бути відсутня G1-фаза. Клітини, які пройшли диференціацію і більше не поділяються, по-
стійно знаходяться у фазі спокою G0. За умов стимуляції мітогенезу (факторами росту, онкогенними вірусами) спочиваючі клітини можуть повер-
нутися до стану, який властивий G1. Якщо такі клітини пройдуть критичну точку, вони вступають до S-фази;
—S-фазу — подвоєння молекул ДНК (реплікацію);
—G2-фазу, яка є кінцевим етапом підготовки клітини до поділу;
—М-фазуклітинного поділу (поділ хромосом, клітин).
Сукупність фаз G1, G0, S та G2 носить назву інтерфази. У клітинному циклі інтерфаза змінюється на суттєво коротшу фазу мітозу.
Біохімічні механізми контролю за вступом клітини до мітозу
Регуляція клітинного циклу здійснюється за допомогою ковалентної модифікації (фосфорилування/дефосфорилування) регуляторних білків, які беруть участь у мітозі, наприклад, вхідного до складу хроматину гістону Н1, ламіну (компо-
нента цитоскелета), факторів транскрипції, білків мітотичного веретена і деяких ферментів.
Фосфорилування цих білків запускає процес
мітозу.
Ключовим білком, який регулює вступ клітини до мітозу (G2/M-перехід), є серин-треонін-про- теїнкіназа, яка включає регуляторну субодиницю
— циклін і каталітичну субодиницю — циклін-
залежну кіназу cdk (від англ. cyclin dependent kinase). Молекула будь-якої cdk складається
тільки з однієї субодиниці, яка сама по собі неактивна. Для активації cdk потрібне зв’язування з нею спеціального білка — цикліну. Термін «циклін» відображає той факт, що концентрація циклінів у клітині протягом клітинного циклу змінюється циклічно.
Після завершення мітозу регуляторна субодиниця циклін взаємодіє з убіквітином — низькомолекулярним пептидом, який є універсальним лігандом і «помічає» білки, які підлягають подальшому розщепленню.
Контроль клітинного циклу
Уході клітинного циклу відбувається само-
контроль власного стану. Контролю піддається стан спадкового матеріалу — хромосом. Залежно від результатів, обирається один із трьох варіантів:
— перехід до наступної стадії циклу;
— більш-менш тривала затримка на поточній стадії — для виправлення виявлених дефектів, якщо це можливо;
— запуск механізму апоптозу, якщо виявлені порушення невиправні.
Унормі протягом ембріогенезу і розвитку відбувається швидка проліферація клітин. Вони можуть втрачати свою властивість контролювати клітинний цикл, що призводить до нерегульованого росту клітин, тобто до раку. Гени, чиї продукти беруть участь у регуляції клітинної проліферації, називають протоонкогенами. Мутація цих генів призводить до підвищення неконтрольованої проліферації клітин, отже, прото-
онкоген може стати онкогеном.
Реплікація ДНК
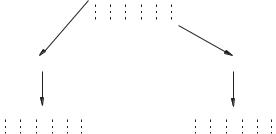
Відповідно до гіпотези Уотсона — Кріка, кожен із ланцюгів подвійної спіралі ДНК є матрицею для реплікації комплементарних дочірніх ланцюгів. Структура подвійної спіралі дозволяє представити простий механізм реплікації ДНК: подвійна спіраль спочатку розкручується, ланцюги розходяться, а потім кожна одноланцюгова половина молекули ДНК добудовується до цілої, дволанцюгової молекули. Наявні нуклеотидні ланцюги є матрицею для синтезу нових ланцюгів; у результаті виходять дві дволанцюгові молекули ДНК, повністю ідентичні вихідній молекулі. Такий спосіб реплікації дістав назву напівконсервативного.
|
G–C–C–A–T–A |
|
C–G–G–T–A–T |
G–C–C–A–T–A |
C–G–G–T–A–T |
G–C–C–A–T–A |
G–C–C–A–T–A |
C–G–G–T–A–T |
C–G–G–T–A–T |
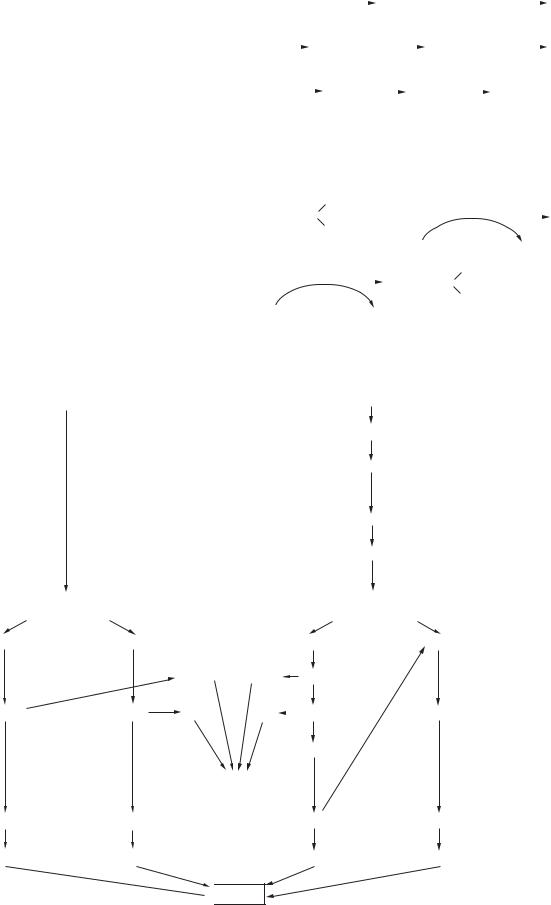
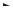
 ДНК
ДНК